|
||||
|
|
Глава 1. ОРГАНЫ РАЗМНОЖЕНИЯ И ОСНОВЫ РЕПРОДУКТИВНОЙ ФИЗИОЛОГИИ СОБАК1.1. ПОЛОВЫЕ ОРГАНЫ САМКИУ самки различают внутренние и наружные половые органы (рис. 1). Внутренние половые органы включают в себя яичники, маточные трубы, матку и влагалище. Яичники (Ovaria, Oophoron) — первичная парная половая железа, выполняющая воспроизводительную и гормональную функции. Яичники овоидной формы, несколько сплюснуты с боков. Во время половой охоты, лютеиновой фазы полового цикла и в период беременности их форма может быть гроздевидной. Размеры яичников у собак сильно варьируют в зависимости от морфо-функционального состояния органа и размеров животного. Например, у собак крупных пород во время лютеиновой фазы полового цикла и в период беременности яичники могут достигать 2–2,5 см в длину и 1…1,5 см в ширину. Яичники располагаются в брюшной полости позади и ниже почек в открытой овариальной бурсе (рис. 2). Стенки овариальной бурсы образованы брыжейками яичников и маточных труб. Брюшное отверстие овариальной бурсы маленькое — не превышает в длину 1…1,5 см. При помощи собственной связки яичник соединен с верхушкой соответствующего рога матки, посредством добавочной связки прикреплен к поясничным позвонкам. Добавочные овариальные связки у собак короткие, содержат много жира и кровеносные сосуды. Указанные анатомические особенности ограничивают доступ к яичникам и затрудняют их оперативное удаление. Снаружи яичник покрыт однослойным кубическим эпителием, под которым находится фиброзная (белочная) оболочка. Паренхима яичника представлена мозговым и корковым веществом. Мозговое вещество состоит из соединительной ткани, сосудов и нервов. В соединительнотканной основе коркового вещества расположены фолликулярный аппарат (первичные, вторичные и третичные фолликулы) и желтые тела (рис. 3). Первичные, или примордиальные, покоящиеся, фолликулы, представляющие собой ооциты первого порядка, окруженные одним слоем фолликулярных клеток, образуются у собак в фетальных (плодных) яичниках. При рождении в яичниках насчитывается 700 000, при наступлении половой зрелости — 250 000, в возрасте 5 лет — 33000, в возрасте 10 лет — 500 первичных фолликулов (McDonald L.E, 1980). 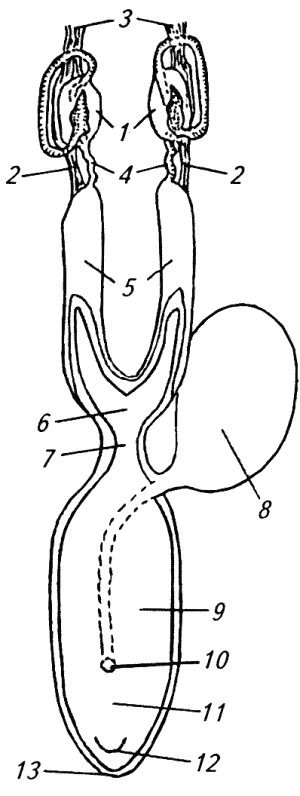 Рис. 1. Половые органы самки, вид сверху: 7 — яичники; 2 — собственные связки яичников; 3 — добавочные связки яичников; 4 — маточные трубы; 5 — рога матки; 6 — тело матки; 7 — шейка матки; 8 — мочевой пузырь; 9 — влагалище; 10 — отверстие уретры; 11 — преддверие влагалища; 12 — клитор; 13 — половые губы Вторичные, или растущие, фолликулы — это ооциты первого порядка, окруженные двумя и более слоями фолликулярных клеток. На данной стадии фолликулогенеза яйцеклетка активно растет и покрывается прозрачной оболочкой. 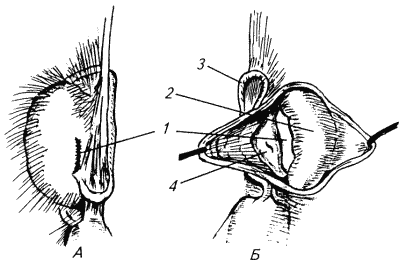 Рис. 2. Овариальная бурса: А — вид сбоку, медиальная поверхность; Б — вид сверху. дорсальная стенка бурсы вскрыта; 1 — брюшное отверстие овариальной бурсы; 2 — яичник; 3 — маточная труба; 4 — воронка маточной трубы. Третичные, или пузырчатые, полостные, граафовы, фолликулы (последняя стадия фолликулогенеза) содержат микро- или макроскопическую полость, заполненную фолликулярной жидкостью. Их стенка выстлана изнутри многослойным фолликулярным эпителием, снаружи — внутренним и наружным слоями соединительнотканной оболочки. Клетки фолликулярного эпителия образуют яйценосный бугорок, в центре которого располагается ооцит первого порядка. Третичные фолликулы вырабатывают эстрогенные гормоны. Гормональная активность граафовых фолликулов зависит от степени их зрелости. Наиболее активны в эндокринном отношении преовуляторные фолликулы, вступившие в финальную стадию своего развития. Незадолго до овуляции они достигают 6…8 мм в диаметре, численность может колебаться от 1 до 14. Овуляция у собак происходит спонтанно. Желтое тело, образующееся на месте овулировавшего фолликула, представляет собой эндокринную железу временной секреции. Клетки желтого тела (лютеоциты) вырабатывают прогестерон — гормон, необходимый для поддержания беременности. Различают желтые тела полового цикла и беременности. У собак желтое тело полового цикла функционирует в течение такого же промежутка времени, что и желтое тело беременности. Маточные трубы (Tuba uterina, salpinx), или яйцепроводы, фаллопиевы трубы, — парный орган в виде извитой трубочки, отходящей от каждого рога матки. Маточные трубы расположены в собственной брыжейке, образованной внутренним листком широкой маточной связки. Их противоположный конец открывается в полость овариальной бурсы; стенка состоит из слизистой, мышечной и серозной оболочек. Слизистая оболочка складчатая, ее однослойный цилиндрический эпителий представлен секреторными и реснитчатыми клетками. В маточных трубах дозревают спермии, оплодотворяется яйцеклетка и зародыш развивается до стадии 16-клеточного бластомера. Половые клетки и зародыш транспортируются в матку благодаря колебаниям ресничек эпителиальных клеток и сокращению гладких мышечных волокон стенки органа. Сократительную активность мышечной стенки маточных труб стимулируют эстрогены и подавляет прогестерон. Матка (Uterus, histera, metra) у собак двурогая, состоит из шейки, тела и рогов. Шейка и тело матки короткие, рога длинные и служат плодовместилищем. Рога расходятся под острым углом, что придает матке форму рогатки. Размер рогов матки у собак сильно варьирует и зависит от размеров животного и физиологического состояния организма — стадии полового цикла и сроков беременности. Стенка матки построена из трех оболочек: наружной — серозной (периметрии), средней — мышечной (миометрий) и внутренней — слизистой (эндометрий). Мышечная оболочка представлена продольными и круговыми слоями, между которыми расположен слой, богатый сосудами и нервами. Сократительную активность миометрия тела и рогов матки стимулируют эстрогены и подавляет прогестерон. Строение слизистой оболочки тела и рогов матки достаточно сложно: она покрыта однослойным цилиндрическим эпителием, в ее толще находятся многочисленные трубчатые железы, протоки которых открываются в полость матки. Железы вырабатывают так называемое маточное молочко, необходимое для питания зародыша. Эндометрий, так же как и миометрий, служит тканью-мишенью для половых гормонов. Эстрогены усиливают васкуляризацию эндометрия, стимулируют рост эндометриальных желез. Чрезмерная васкуляризация эндометрия приводит к просачиванию (диапедезу) клеток крови в просвет матки и появлению в стадию проэструса геморрагических выделений из половой щели. Прогестерон вызывает ветвление трубчатых желез и стимулирует выработку маточного молочка. Во время беременности у собак, так же как и у других плацентарных животных, из слизистой оболочки матки и сосудистой оболочки плода образуется плацента, которая по микроскопическому строению принадлежит к типу эндотелиохориальных, по макроскопическому — к типу зональных. Во время родов отпадает только детская часть плаценты. Шейка матки (Cervix uteri) имеет узкий канал, толстую стенку с хорошо развитым мышечным слоем. У собак шейка матки достигает в длину 1… 1,5 см и характеризуется отсутствием четких границ с телом матки и влагалищем. Вход в цервикальный канал со стороны влагалища прикрыт постцервикальной влагалищной складкой и недоступен для вагинального осмотра. Шейка выполняет функцию сфинктера матки. Полное раскрытие ее канала и постцервикальной влагалищной складки (ложной шейки матки) отмечают во время родов, частичное — во время течки, половой охоты и в послеродовом периоде. Раскрытие шейки матки при родах стимулируют эстрогены и релаксин, во время течки и половой охоты — только эстрогенные гормоны. Эпителий слизистой оболочки шейки матки однослойный цилиндрический и представлен главным образом секреторными клетками, вырабатывающими слизистый секрет с бактерицидными и бактериостатическими свойствами. Матка расположена в брюшной полости, ее поддерживают широкие и круглые маточные связки. Широкие связки матки — это двойные листки брюшины, идущие от малой кривизны рогов, боковой поверхности тела, шейки матки и краниальной части влагалища к боковым стенкам таза. Круглые связки матки в виде шнуров начинаются у верхушки рогов матки и заканчиваются у внутреннего отверстия пахового канала. 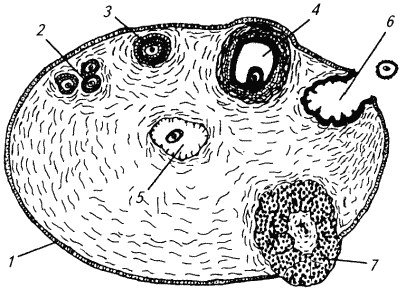 Рис 3. Схематическое изображение яичника, саггитальный срез: 1 — покровный эпителий; 2 — первичные фолликулы; 3 — вторичный фолликул; 4 — третичный фолликул; 5 — атрезия фолликула; 6 — овулировавший фолликул; 7 — желтое тело Влагалище (Vagina), или вагина, находится в тазовой полости между шейкой матки и отверстием уретры (мочеиспускательный канал). Представляет собой тонкостенную эластическую трубку и служит органом совокупления и родовым каналом. Изнутри стенка влагалища выстлана слизистой оболочкой, лишенной желез и покрытой многослойным плоским эпителием. Под влиянием эстрогенных гормонов в период проэструса и особенно эструса (половой охоты) возрастает число слоев эпителиальных клеток, поверхностные клетки ороговевают, лишаются ядра, в их цитоплазме накапливается кератин. Под слизистой оболочкой расположены два слоя мышц: продольный и циркулярный (поперечный). Краниальная часть влагалищной трубки снаружи покрыта серозной (брюшинной) оболочкой, остальная же ее часть — рыхлой соединительной тканью, которая вместе с параректальной соединительнотканной клетчаткой обеспечивает фиксацию вагины и прямой кишки в тазовой полости. Наружные половые органы включают в себя преддверие влагалища, половые губы и клитор. Преддверие влагалища (Vestibulum vaginae) служит мочеполовым каналом. Его слизистая оболочка не содержит вестибулярных желез, покрыта многослойным плоским эпителием и выполняет только защитную функцию. Мышечная оболочка хорошо развита и образует сфинктер преддверия влагалища, обеспечивающий сцепление половых органов самки и самца во время полового акта. Границей между влагалищем и его преддверием служит отверстие уретры. Девственная плева (Hymen) у собак плохо развита или отсутствует. Преддверие влагалища каудально переходит в половую щель (Rima pudendi), ограниченную половыми губами (Labia vulvae), или вульвой, половой петлей. Верхний угол вульвы закруглен, нижний заострен. В нижнем углу половой щели располагается клитор (Clitoris) — гомолог полового члена, не содержащий половой косточки. Клитор состоит из фиброзной, жировой и эректильной тканей и богат сенсорными нервными окончаниями. Половые органы самок кровоснабжаются сосудами, отходящими от овариалъной, или яичниковой, артерии (Arteria ovaricd) и ветвями внутренней срамной артерии (A. pudenda inlerna). Овариальная артерия ответвляется непосредственно от аорты позади почечной артерии и разделяется на две ветви — трубную (Ramus tubarius) и маточную (R. uterinus), которые васкуляризируют яичники, маточные трубы и краниальную часть рогов матки. Внутренняя срамная артерия берет начало от внутренней подвздошной артерии (A. iliaca intema) и разделяется на несколько ветвей. В кровоснабжении половых органов самок основное значение имеют две из них — влагалищная (A. vaginalis) и вентральная промежностная (A. perinealis ventralis) артерии. Влагалищная артерия питает стенку влагалища и на уровне шейки матки переходит в маточную артерию (A. uterina), которая васкуляризирует стенки шейки, тела и 2/3 рогов матки. Ветви вентральной промежностной артерии питают наружные половые органы и ткани промежности. Овариальные вены (Venae ovaricae) служат главным стволом, по которому венозная кровь отводится из генитальных органов. При этом правая овариальная вена (Vena ovarica dextra) впадает в заднюю полую вену (V. cava caudalis), левая (V. ovarica sinistra) — в почечную вену (V. renalis). Лимфатическая система половых органов самок очень хорошо развита. Лимфа собирается в региональные лимфатические узлы — тазовые, крестцовые и паховые, выполняющие фильтрационно-барьерную и иммунную функции. Важнейшие функции половых органов самок: В иннервации органов размножения самок участвуют симпатическая и парасимпатическая системы. Симпатические волокна отходят от тазового сплетения (Plexus pelvinus), парасимпатические — от крестцовых нервов (Nervi sacrales). Наружные половые органы и влагалище также хорошо снабжены сенсорными нервными волокнами. 1.2. ПОЛОВЫЕ ОРГАНЫ САМЦАОрганы размножения самца состоят из семенников, их выводных протоков (придатки семенников, спермиопроводы и мочеполовой канал), предстательной железы, полового члена, препуция и мошонки (рис. 4). 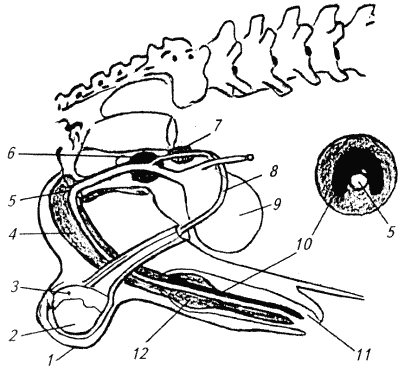 Рис. 4. Половые органы самца, вид сбоку: 1 — мошонка; 2 — семенник; 3 — придаток семенника; 4 — половой член; 5 — мочеполовой канал; 6 — предстательная железа; 7 — ампула спермиопровода; 8 — спермиопровод; 9 — мочевой пузырь; 10 — половая косточка; 11 — препуций; 12 — луковица головки полового Семенники (Testis, orchis, didymis), или тестикулы, — первичный парный половой орган, выполняющий воспроизводительную и гормональную функции: вырабатывает мужские половые клетки спермии и мужской половой гормон тестостерон. Семенники овальной формы, плотноэластической консистенции и достигают в длину 2…4 см. На семеннике различают головчатый и хвостатый концы, свободный и придатковый края, латеральную и медиальную поверхности. Снаружи семенник покрыт собственной влагалищной (серозной) оболочкой, под которой располагается белочная оболочка. Ее радиальные тяжи делят паренхиму органа на многочисленные пирамидальные дольки и формируют соединительнотканное средостение семенника. Вершина пирамидальных долек обращена к средостению семенника, основание — к белочной оболочке. Каждая долька содержит несколько извитых канальцев, окруженных рыхлой соединительной тканью с большим количеством кровеносных сосудов. В соединительнотканной основе пирамидальных долек находятся клетки Лейдига, вырабатывающие андрогенный гормон тестостерон. Извитые канальцы начинаются слепым мешком и, сливаясь у верхушки пирамидальной дольки, впадают в прямые канальцы семенников, протоки которых открываются в сеть семенника (рис. 5). Спермии образуются в извитых канальцах семенников, функция прямых канальцев и сети семенника — транспорт половых клеток. Стенка извитых канальцев состоит из двух слоев: соединительнотканного и эпителиального, отделенных друг от друга базальной мембраной, которая служит гематотестикулярным барьером. 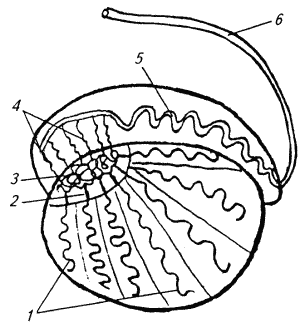 Рис. 5. Схематическое изображение семенника и его придатка, сагиттальный срез: 1 — извитые канальцы; 2 — прямые канальцы; 3 — сеть семенника; 4 — спермиовыносящие канальцы; 5 — канал придатка семенника; 6 — спермиопровод 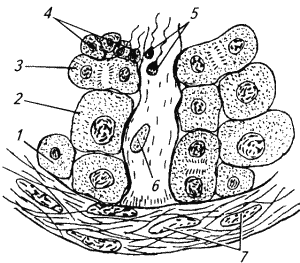 Рис. 6. Микроструктура стенки извитого канальца семенника: 1 — сперматогоний; 2 — сперматоцит первого порядка; 3 — сперматоцит второго порядка; 4 — сперматиды; 5 — спермии; 6 — клетка Сертоли; 7 — фиброциты Процесс образования спермиев характеризуется четким временным циклом и продолжается на протяжении всей репродуктивной жизни самца. Сперматогенный эпителий половозрелых собак многослойный и состоит из сперматогоний, сперматоцитов первого и второго порядков, сперматид и спермиев (рис. 6). Все эти клетки соединены между собой синцитиальными отростками клеток Сертоли, которые выполняют питательную и секреторную функции: продуцируют тестикулярную жидкость, вырабатывают белок, связывающий тестостерон, и гормон ингибин, тормозящий секрецию фолликулостимулирующего гормона (ФСГ). Мошонка (Scrotum) — особое образование брюшной стенки, в котором располагаются семенники. Выполняет защитную и терморегулирующую функции. У собак мошонка находится между бедер и представляет собой кожно-мышечный мешок, разделенный перегородкой на правую и левую камеры, которые через соответствующие паховые каналы сообщаются с брюшной полостью. Кожа мошонки у собак — с редким волосяным покровом, содержит большое количество сальных и потовых желез. За счет потовых желез мошонка способна активно поддерживать оптимальную для сперматогенеза температуру в семенниках — на несколько градусов Цельсия ниже температуры тела животного. Секрет сальных желез уменьшает теплоотдачу и защищает кожу мошонки от неблагоприятных факторов внешней среды. Кожа тесно сращена с мышечно-эластической оболочкой, образующей перегородку мошонки. За мышечно-эластической располагается общевлагалищная оболочка семенника, представляющая собой париетальный листок брюшины. Мышечно-эластическая и общевлагалищная оболочки рыхло соединены между собой, их легко отделить друг от друга. Общевлагалищная оболочка через влагалищную (тестикулярную) связку, переходящую на хвостатый конец семенника, соединена с собственной влагалищной оболочкой семенника. К наружной поверхности общевлагалищной оболочки сбоку и сзади крепится подниматель семенника (М. cremaster), который с мышечно-эластической оболочкой участвует в регулировании температуры в семенниках и его придатках, изменяя объем мошонки и расстояние между семенниками и паховыми каналами. Семенники у собак располагаются в полости мошонки почти в горизонтальном положении. Они подвешены спереди на семенном канатике, сзади — на собственной связке семенника. Семенной канатик (Funiculus spermaticus) представляет собой тяж, простирающийся от головчатого конца семенника до внутреннего пахового кольца. Состоит из поднимателя семенника, сильно извитых тестикулярных сосудов, нервов и спермиопровода. Густая сеть венозных сосудов, обеспечивающих понижение температуры артериальной крови в семенниках, образует венозное сплетение. Придатки семенников (Epididymis) — парный орган, тесно примыкающий к поверхности семенников. В придатке семенника различают головку, тело и хвост. Головка состоит из 12…18 спермиовыносящих канальцев, соединяющих сеть семенника с сильно извитым каналом придатка семенника, от которого начинается спермиопровод. В придатке семенника спермии созревают и концентрируются. В функции органа входят также хранение и транспорт спермиев. По мере продвижения по каналу придатка семенника спермии освобождаются от цитоплазматической капли (остатков цитоплазмы сперматиды), покрываются защитной оболочкой, приобретают отрицательный электрический заряд, способность к прямолинейно-поступательному движению и оплодотворению. В кислой бескислородной среде при температуре ниже температуры тела животного на несколько градусов Цельсия они сохраняют оплодотворяющую способность в течение нескольких месяцев. Спермиопроводы (Ductus deferens) — парный трубчатый орган, состоящий из слизистой, мышечной и серозной оболочек; обеспечивает транспорт спермиев из канала хвоста придатка семенника в мочеполовой канал. В спермиопроводе различают четыре части: семенниковую, соответствующую длине семенника; канатиковую, проходящую в составе семенного канатика до поверхностного пахового кольца; паховую — в паховом канале; тазовую часть — участок от глубокого пахового кольца до места впадения в мочевой канал. Около шейки мочевого пузыря конечные части спермиопроводов расширяются, становятся веретенообразными и образуют ампулы. Стенка ампул содержит секреторно активные трубчатые железы. Мочеполовой канал (Canalis urogenitalis), обеспечивающий транспорт мочи и спермы, начинается с места впадения в мочевой канал спермиопроводов. В нем различают тазовую (до седалищной вырезки) и пенисную части. Слизистая оболочка мочеполового канала у собак не содержит уретральных желез и представлена многослойным плоским неороговевающим эпителием. За слизистой оболочкой находится слой гладких мышечных волокон. Пенисная часть мочеполового канала окружена губчатой тканью и располагается в специальном желобе половой косточки. Мочеполовой канал заканчивается на головке полового члена мочеполовым отверстием. Предстательная железа (Prostata) у собак двудольчатая, по строению трубчато-альвеолярная. Располагается в тазовой полости над шейкой мочевого пузыря, протоки открываются в тазовую часть мочеполового канала. Предстательная железа вырабатывает секрет, входящий в состав спермы. Пузырьковидные и луковичные железы у собак отсутствуют. Половой член (Penis), или пенис, — орган совокупления и мочевыделения. У собак он васкулярного типа с половой косточкой (Os penis), придающей ему упругость. В пенисе различают корень, тело и головку. Корень состоит из двух ножек, берущих начало от седалищных бугров. Ножки, окруженные развитой луковично-кавернозной мышцей (М. bulbospongiosus), соединяются над мочеполовым каналом и вместе с ним образуют тело пениса, заканчивающееся головкой. Половая косточка, находящаяся в головке полового члена, на 2/3 заполняет мочеполовой канал, сужая его отверстие. У собак крупных пород половая косточка достигает 8… 10 см в длину. Основа полового члена — два кавернозных тела и одно губчатое, окружающее мочеполовой канал и образующее у собак луковицу полового члена. Эти тела покрыты белочными оболочками и содержат многочисленные сообщающиеся между собой полости (каверны), способные при сокращении луковично-кавернозной (эректорной) мышцы во время полового возбуждения накапливать кровь и вызывать эрекцию полового члена. Сперма из полового члена выделяется благодаря перистальтическим сокращениям стенки мочеполового канала и ритмичным сокращениям луковично-кавернозной мышцы, расположенной у основания пениса. Коаниальная часть пениса находится в препуциальном мешке на вентральной поверхности живота. Снаружи препуций покрыт кожей изнутри выстлан многослойным плоским неороговевающим эпителием (париетальный листок), одевающим также головку полового члена (висцеральный листок). Париетальный листок поепуция у собак не содержит препуциальных желез. В препуциальном мешке половой член удерживается специальной ретрактооной мышцей (М. retractor penis), состоящей из гладких мышечных волокон. Мышца берет начало у первых хвостовых позвонков и заканчивается у основания головки пениса. При эрекции половой член увеличивается в размере и выходит за пределы препуциального мешка. Луковица пениса сильно набухает, что способствует сцеплению половых органов самца и самки во время полового акта. Половые органы самцов кровоснабжаются семенной артерией (A. testicularis) и ветвями внутренней срамной артерии. Семенная артерия отходит от аорты и питает семенник и его придатки. Внутренняя срамная артерия берет начало от внутренней подвздошной артерии и дает три основные ветви, участвующие в кровоснабжении половых органов самцов: предстательную (A. prostatica), вентральную промежностную и артерию пениса (A. penis). Предстательная артерия васкуляризирует простату и мочевой пузырь. Вентральная промежностная — ткани промежности и мошонки. Артерия пениса разделяется на три ветви — дорсальную артерию полового члена (A. dorsalis penis), артерию луковицы пениса (A. buibi penis) и глубокую артерию пениса (A. profunda penis). Отток крови из половых органов обеспечивают одноименные вены. Лимфа из генитальных органов собирается в региональные лимфатические узлы. В иннервации половых органов самцов также принимают участие вегетативная и соматическая нервная системы. Наружные половые органы-мошонка, препуций и особенно краниальная часть полового члена — хорошо снабжены сенсорными нервными окончаниями. Раздражение во время полового акта термо- и баро-рецепторов головки полового члена инициирует эякуляцию (выделение спермы). Барорецепторам принадлежит ведущая роль в проявлении рефлекса эякуляции. Важнейшие функции половых органов самцов суммированы ниже. 1.3. РАЗВИТИЕ ПОЛОВЫХ ОРГАНОВ И ОСОБЕННОСТИ ОВО- И СПЕРМАТОГЕНЕЗАВ процессе эмбрионального развития у индивидуума одновременно закладываются мужские и женские половые органы. Индифферентная половая система состоит из первичных гонад, мезонефральных (вольфовых) и парамезонефральных (мюллеровых) протоков, мочеполового синуса, полового бугорка и генитальных складок. Особенности дифференцировки половых органов плодов у собак показаны в таблице 1 и на рисунке 7. 1. Особенности внутриутробного развития половых органов у собак Половые железы закладываются на внутренней поверхности первичной почки. Первичная гонада состоит из клеток целомического эпителия (наружный корковый слой), мезенхимы (внутренний мозговой слой) и первичных половых клеток внегонадального происхождения — гоноцитов, которые мигрируют в индифферентную половую железу из эндодермы желточного мешка. Половая дифференцировка гонад индуцируется набором половых хромосом, образующихся в зиготе при слиянии спермия с яйцеклеткой. Половые клетки в отличие от соматических содержат гаплоидный набор хромосом. Спермий может нести либо Х-, либо Y-хромосому, яйцеклетка несет только Х-хромосому. Набор половых хромосом XY индуцирует дифференцировку гонад по мужскому типу, набор XX — по женскому типу. При развитии гонад по мужскому типу гоноциты локализуются во внутреннем мозговом слое половой железы. Они внедряются в семенные тяжи, образованные клетками целомического эпителия. Семенные тяжи дифференцируются на сеть семенника, прямые и извитые канальцы семенников. В извитых канальцах гоноциты трансформируются в сперматогонии, клетки целомического эпителия — в клетки Сертоли. В это же время из клеток мезенхимы образуются клетки Лейдига. Фетальные тестикулы гормонально активны. Клетки Сертоли вырабатывают антимюллеров фактор, вызывающий регрессию парамезонефральных каналов, клетки Лейдига — тестостерон, обеспечивающий развитие из эмбриональных закладок вторичных половых органов самца: придатков семенников, спермиопроводов, предстательной железы, полового члена, препуция и мошонки. 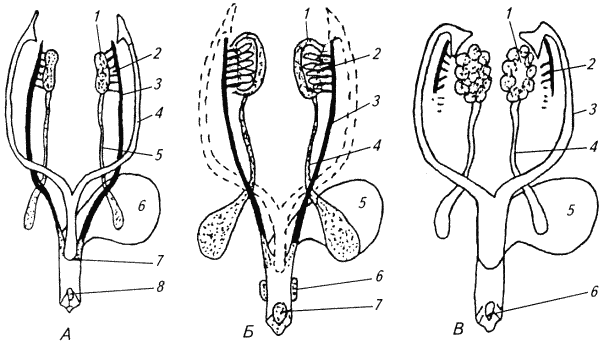 Рис. 7. Дифференцировка половых органов: А — индифферентная стадия: 1 — гонада; 2 — первичная почка (мезонефрос); 3 — мезонефральные (вольфовы) протоки; 4 — парамезонефральные (мюллеровы) протоки; 5 — паховый тяж; 6 — мочевой пузырь; 7 — мочеполовой синус; 8 — половой бугорок; Б — формирование половых органов самца: 1 — семенник; 2 — придаток семенника; 3 — спермиопровод; 4 — тестикулярная связка (паховый тяж); 5 — мочевой пузырь; 6 — предстательная железа; 7 — половой член; В — формирование половых органов самки: 1 — яичник; 2 — остатки первичной почки (пара- и эпофорон); 3 — маточные трубы; 4 — круглая маточная связка (паховый тяж); 5 — мочевой пузырь; 6 — клитор В конце фетального периода развития семенники оказываются в паховом канале и на 10… 14-й день после рождения щенка опускаются в мошонку за счет дифференцированного роста поддерживающих связок семенника и прежде всего пахового тяжа тестикулярной связки. Отсутствие семенников в мошонке может быть обусловлено пороками развития половых желез — крипторхиз-мом, анорхизмом и эктопией. В постнатальном периоде созревают половая и гипоталамо-гипофизарная системы, устанавливается взаимодействие их гормонов, развиваются вторичные половые признаки (время полового созревания). Сперматогенез — процесс образования и созревания мужских половых клеток, предшествует наступлению половой зрелости и продолжается на протяжении всей репродуктивной жизни самца. В среднем продолжительность сперматогенеза у собак составляет 56,4 дня. Спермии образуются в извитых канальцах семенников. В ходе сперматогенеза диплоидные сперматогонии трансформируются в гаплоидные дифференцированные мужские клетки — спермии. Родоначальные половые клетки делятся по типу митоза и мейоза. По типу митоза размножаются сперматогонии. В процессе каждого митотического деления сперматогонии дифференцируются на активные, промежуточные и неактивные варианты. Сперматоциты первого порядка, образующиеся из активных сперматогониев, растут и вступают в первое мейотическое деление, при котором из одного сперматоцита первого порядка образуются два сперматоцита второго порядка. Во время первого мейотического деления происходит кроссинговер — обмен блоками генов внутри одной хромосомы и между гомологичными хромосомами, что создает возможность наследственного варьирования в потомстве. После непродолжительного периода покоя сперматоциты второго порядка вступают во второе мейотическое деление, в результате которого образуются четыре сперматиды с гаплоидным набором хромосом. Сперматиды больше не делятся, а видоизменяются, что приводит к образованию спермиев. Оогенез — процесс образования женских половых клеток. Внутриутробное развитие репродуктивной системы у плодов самок начинается позднее, чем у плодов самцов. Гоноциты, локализующиеся в наружном корковом слое гонад, превращаются в оогонии, которые, так же как сперматогонии, содержат диплоидный набор хромосом. Оогонии интенсивно размножаются путем митотического деления. Завершив последнее митотическое деление, оогонии вступают в первую стадию мейоза и превращаются в ооцит первого порядка. Созревание ооцита первого порядка приостанавливается на стадии диктиотены профазы первого мейоза. Блокада мейоза совпадает по времени с процессом образования первичного фолликула — формированием вокруг ооцита первого порядка одного слоя фолликулярных клеток. К моменту рождения плода яичники морфологически сформированы. В корковом слое находятся множественные первичные фолликулы. Мозговой слой (остатки мезенхимального слоя индифферентной гонады) состоит из соединительной ткани, сосудов и нервов. В отличие от фетальных семенников гормональная активность яичников плода незначительна. Гормоны яичников не оказывают решающего влияния на половую дифференцировку репродуктивной системы самок в период преднатального развития. При врожденном отсутствии гонад или в результате их удаления на стадии индифферентной гонады развиваются половые органы только женского типа. При гермафродитизме (аномалии полового развития, когда гонады дифференцируются сразу в двух направлениях — овариальном и тестикулярном) развитие внутренних и наружных гениталий зависит от наличия и активности тестостерон-секретирующих клеток в гонаде смешанного типа. В постнатальном периоде во время полового созревания формируется суточная цикличность и увеличивается выделение гонадотропинов, под влиянием которых возрастает секреция эстрогенных гормонов яичников, устанавливается взаимодействие между яичниками и гипоталамо-гипофизарной системой. Фолликулогенез носит незавершенный характер. Фолликулы дегенерируют на разных стадиях своего развития. Процесс дегенерации третичных фолликулов называется атрезией. Генеративную активность яичники начинают проявлять при наступлении половой зрелости. Созревание половых клеток носит циклический характер: в течение каждого полового цикла несколько третичных фолликулов вступают в финальную стадию своего развития, созревают и овулируют. Яйцеклетка овулирует на стадии ооцита первого порядка (не содержит полярных телец). Она правильной шарообразной формы, ее диаметр 1,2?10-5 мм, а вместе с прозрачной оболочкой 1,56?10-5 мм (Hoist P.A. et al., 1971). Ооцит созревает в верхней трети яйцепроводов, претерпевая два мейотических деления. Первое мейотическое деление инициируется преовуляторным выбросом лютеинизирующего гормона (ЛГ) и приводит к образованию ооцита второго порядка и отделению в перивителлиновое пространство первого полярного тельца, содержащего незначительное количество цитоплазмы и лишние хромосомы. При проникновении в яйцеклетку спермия ооцит второго порядка претерпевает второе деление мейоза, в результате которого образуются одна зрелая яйцеклетка с гаплоидным набором хромосом, способная к оплодотворению, и второе полярное тельце. 1.4. НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ ПОЛОВЫХ ПРОЦЕССОВВажнейшая функция нервной системы — управление деятельностью целостного организма при помощи нервных и гуморальных сигналов на основании сбора, анализа и интеграции информации, исходящей от отдельных частей организма и из окружающей среды. По топографическому признаку нервную систему подразделяют на центральную и периферическую. Центральная нервная система (ЦНС) включает в себя спинной и головной мозг, периферическая — спинномозговые и черепные нервы, их ветви и сплетения. 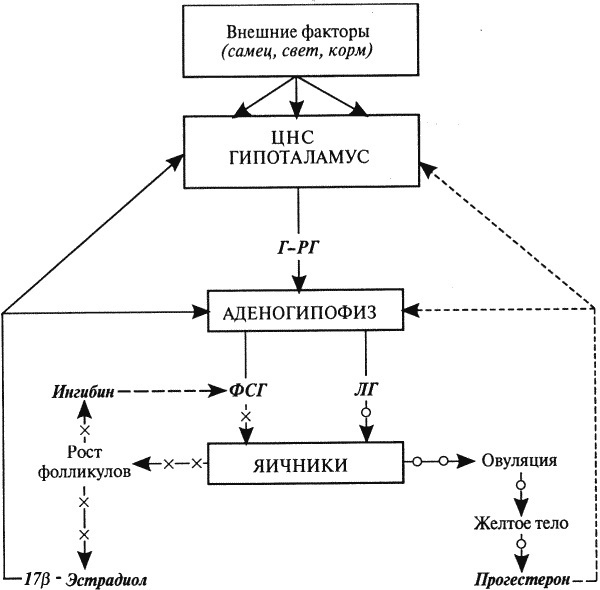 Рис. 8. Схема нейроэндокринной регуляции половых процессов у самок Рис. 8. Схема нейроэндокринной регуляции половых процессов у самок [Сплошными линиями показана прямая и положительная обратная связь (стимуляция), прерывистыми — отрицательная (блокирование)]: гормон гипоталамуса Г-РГ стимулирует выделение из аденогипофиза ФСГ и ЛГ; ФСГ активирует рост и развитие фолликулов и выработку ими эстрогенов и ингибина — ингибин избирательно блокирует секрецию ФСГ; эстрогены, воздействуя на органы-мишени (ЦНС, вторичные половые органы), индуцируют проявление течки, полового возбуждения и охоты; в конце проэструса и в начале эструса, когда концентрация прогестерона низкая, преовуляторный пик Пр-эстрадиола инициирует циклический выброс Г-РГ, ФСГ и ЛГ; преовуляторный пик ЛГ индуцирует созревание преовуляторных фолликулов, их овуляцию, базальная секреция ЛГ — формирование желтых тел и выработку ими прогестерона; прогестерон через механизмы отрицательной обратной связи контролирует секрецию Г-РГ, ФСГ и ЛГ: высокие его концентрации блокируют, а низкие стимулируют выделение этих гормонов По функциональному признаку различают соматическую и вегетативную нервную систему. Соматическая система иннервирует органы тела (сому) и связывает организм с внешней средой при помощи органов чувств, кожной чувствительности и движения. Центры соматической системы находятся в ЦНС, высший отдел которой — кора головного мозга — контролирует высшую нервную деятельность. Вегетативная, или автономная, система, обеспечивающая иннервацию органов и систем организма, в состав которых входят гладкие мышечные волокна и железистый эпителий (органы пищеварения, дыхания, кровоснабжения, выделения, размножения и внутренней секреции), включает в себя симпатическую и парасимпатическую части. Центры симпатической нервной системы расположены в грудопоясничном отделе спинного мозга, парасимпатической — в стволе головного мозга и крестцовом отделе спинного мозга. Нервный импульс в периферических синапсах симпатической системы передается при помощи норадреналина, парасимпатической — ацетилхолина. Симпатическая и парасимпатическая части координируют работу внутренних органов, оказывая на них противоположное действие, например, у самок возбуждение b-адренорецептеров симпатической системы способствует расслаблению матки, их блокада или же возбуждение холинорецептеров парасимпатической системы, наоборот, стимулирует сокращение органа. У самцов симпатическая часть стимулирует рефлекс эякуляции, парасимпатическая — эрекции. Проявление полового цикла и половых рефлексов зависит от взаимодействия нервной и эндокринной систем. Схема нейроэндокринной регуляции половых процессов у самок показана на рисунке 8, половых процессов у самцов — на рисунке 9. Гипоталамус — надбугорная область промежуточного мозга, является одновременно нервным образованием и эндокринной железой. Вырабатывает окситоцин, вазопрессин и еще 10 гипофизотропных нейрогормонов, семь из которых оказывают на переднюю долю гипофиза стимулирующее действие (либерины), три — ингибирующее (статины). В регуляции воспроизводительной функции принимают участие окситоцин, пролактостатин, гонадолиберин и кортиколиберин. 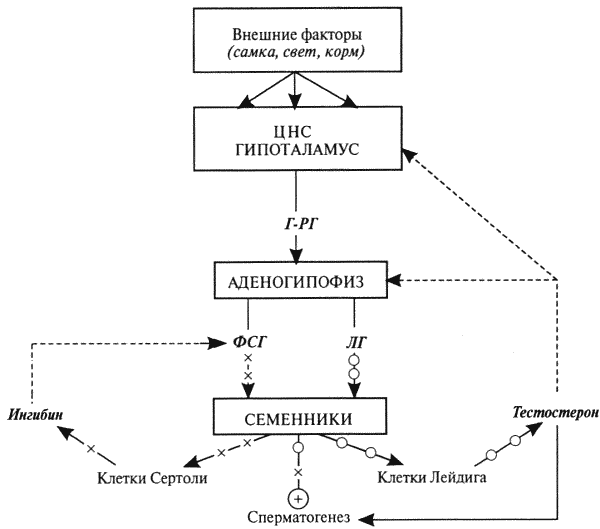 Рис. 9. Схема нейроэндокринной регуляции половых процессов у самцов Рис. 9. Схема нейроэндокринной регуляции половых процессов у самцов [Сплошными линиями показана прямая связь (стимуляция), прерывистыми — отрицательная обратная связь (блокирование)]: гормон гипоталамуса Г-РГ стимулирует выделение из аденогипофиза ФСГ и ЛГ; ЛГ стимулирует выработку клетками Лейдига гормона тестостерона; тестостерон поддерживает сперматогенез, половое влечение и через механизмы отрицательной обратной связи контролирует секрецию Г-РГ, ФСГ и ЛГ: высокие его концентрации блокируют, а низкие — стимулируют выделение этих гормонов; ФСГ стимулирует выработку клетками Сертоли гормона ингибина; инги-бин через механизмы обратной связи подавляет секрецию ФСГ Окситоцин представляет собой нанопептид; вырабатывается гипоталамусом и аккумулируется в задней доле гипофиза (нейро-гипофизе). Окситоцин стимулирует сократительную активность матки, принимает участие в родовом акте и способствует отдаче секрета молочных желез во время акта сосания. У самцов окситоцин, по-видимому, вызывает перистальтические сокращения в спермиопроводах. Секреция гормона в организме регулируется нейрорефлекторным путем (рис. 10). Пролактостатин, или пролактинингибирующий фактор (ПИФ), блокирует секрецию пролактина передней долей гипофиза (аденогипофизом). Секрецию пролактостатина стимулирует дофамин. На применении агонистов дофамина основано подавление лактации в ветеринарной и медицинской практике. Гонадолиберин, или люлиберин, гонадотропин-рилизинг-фактор, гонадотропин-рилизинг-гормон Г-РГ, ЛГ-РГ, ФСГ/ЛГ-РГ, — декапептид, регулирующий синтез и секрецию гонадотропных гормонов гипофиза — фоллитропина (фолликулостимулирующий гормон, или ФСГ) и лютропина (лютеинизирующий гормон, или ЛГ). ФСГ и ЛГ по химическому составу представляют собой глю-копротеиды. ФСГ стимулирует у самок рост и развитие фолликулов, у самцов — сперматогенез; ЛГ — созревание преовуляторных фолликулов, их овуляцию, образование желтого тела и выработку прогестерона. Вместе с ФСГ лютропин инициирует также секрецию эстрогенов третичными фолликулами. У самцов ЛГ стимулирует продукцию клетками Лейдига мужского полового гормона тестостерона. 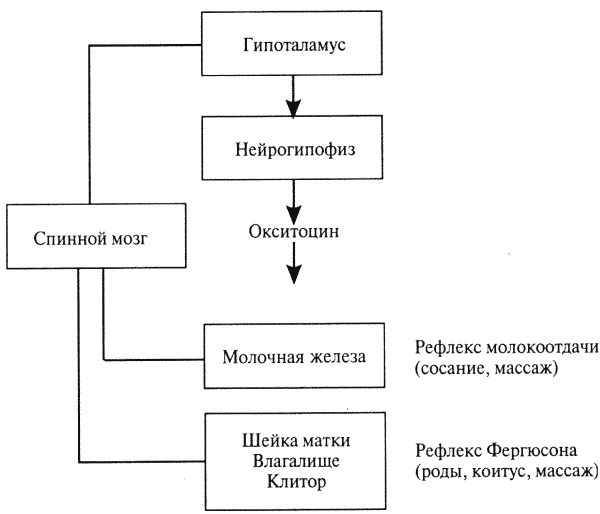 Рис. 10. Нейрорефлекторные пути высвобождения окситоцина гипоталамо-гипофизарной системой Рис. 10. Нейрорефлекторные пути высвобождения окситоцина гипоталамо-гипофизарной системой Пролактин (ПРЛ), лактогенный или лютеотропный гормон — полипептид, вырабатываемый аденогипофизом; у самок стимулирует процесс образования молока, поддерживает лактацию и во второй половине беременности проявляет лютеотропные свойства. У самцов его действие неизвестно. Характерная особенность ПРЛ — органы-мишени пролактина (молочная железа, желтое тело) не синтезируют гормоны, тормозящие его секрецию (отсутствие обратной связи). Существует два типа секреции гонадотропинов: тоническая и циклическая. Тоническая секреция непрерывна, ее регистрируют у самцов и самок на протяжении всей жизни. Циклическая секреция ФСГ и ЛГ предшествует овуляции, ее регистрируют у половозрелых самок. Преовуляторный выброс ЛГ у спонтанно овули-рующих животных инициируется преовуляторным пиком эстрадиола, у рефлекторно овулирующих (кошки, кролики, верблюды) — половым актом. Эпифиз, или шишковидная железа, — верхний мозговой придаток, относящийся к структурам промежуточного мозга. Вырабатывает нейросекрет мелатонин, характеризующийся (в зависимости от вида животного) анти- и прогонадальной активностью: способностью тормозить или, наоборот, стимулировать секрецию гонадолиберина. Отличительная особенность эпифиза — зависимость его секреторной активности от освещенности (фотопериодизм). Активность железы возрастает в темное время суток. Нейрогормон эпифиза контролирует суточный гормональный ритм в организме. У многих моно- (волки, койоты, шакалы, дикие собаки Динго и др.) и полициклических животных (представители семейства кошачьих, овцы, лошади и др.) световой фактор служит основным климатическим регулятором сезонности размножения. Роль мелатонина у собак остается невыясненной, так как собаки проявляют половую активность вне зависимости от сезона года. Физиологическая роль половых гормонов в организме самок и самцов исключительно многообразна. Эндокринные части яичников продуцируют эстрогены, прогестерон и ингибин, семенников — тестостерон и ингибин. Эстрогены — половые гормоны стероидной природы, состоят из 18 атомов углерода (С18). Вырабатываются растущими и созревающими третичными фолликулами и плацентой. Различают три фракции эстрогенов — эстрадиол, эстрон и эстриол. Эстрадиол, наиболее активный из перечисленных, представляет собой первичный эстроген, который может превращаться в эстрон и эстриол. В организме самок эстрогены ответственны за развитие вторичных половых признаков и выводных протоков молочных желез. Эстрогены индуцируют течку, половое возбуждение и охоту, вызывают пролиферацию эндометрия, миометрия, эпителия слизистой оболочки влагалища и его преддверия, усиливают кровоснабжение половых органов, способствуют раскрытию цервикального канала, стимулируют сократительную активность матки, маточных труб и принимают активное участие в родовой деятельности, через механизмы положительной обратной связи индуцируют Преовуляторный выброс ЛГ у спонтанно овулирующих животных. Прогестерон — стероидный гормон (C21). Вырабатывается желтым телом полового цикла и беременности, а также плацентой. У собак желтое тело — основной продуцент прогестерона на протяжении всего периода беременности. Овариоэктомия приводит к прерыванию беременности на любой ее стадии. Прогестерон переводит эндометрий в секреторно активное состояние, подготавливает его к прикреплению зародыша, поддерживает в матке условия, необходимые для развития эмбриона и плода, блокирует сократительную активность матки, вызывает закрытие цервикально-го канала, подавляет созревание фолликулов, проявление течки, полового возбуждения и охоты, стимулирует развитие альвеол молочной железы и тормозит секрецию ЛГ. 2. Участие гормонов в регулировании репродуктивной функции у собак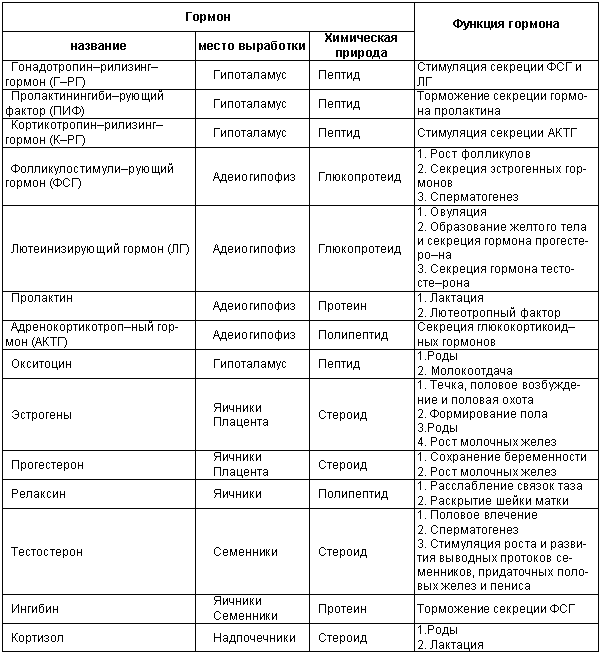 Ингибин — пептидный гормон, вырабатываемый фолликулярным эпителием третичных фолликулов и клетками Сертоли. Тормозит секрецию ФСГ в организме самок и самцов. Релаксин — пептидный гормон, вырабатываемый желтым телом в конце беременности. Подготавливает материнский организм к родовому акту, вызывая расслабление связок таза, мышц шейки матки, влагалища и его преддверия. Тестостерон — мужской половой гормон стероидной природы (С19), вырабатываемый в семенниках, яичниках, коре надпочечников. В семенниках тестостерон синтезируется в клетках Лейдига, в яичниках — в третичных фолликулах, являясь лишь промежуточным этапом биосинтеза. Физиологическая роль тестостерона в эмбриональный период — участие в половой дифференцировке организма. Тестостерон необходим для развития вторичных половых признаков, поддержания сперматогенеза, стимуляции полового влечения и регуляции секреции ЛГ у самцов. В регуляции половых процессов принимают также участие материнские и фетальные глюкокортикостероиды и простагландин Фзц. Глюкокортикоиды — стероидные гормоны, вырабатываемые надпочечниками матери и плода. Кортизол (гормон стресса) — наиболее активный и принципиальный глюкокортикоидный гормон, принимающий участие в регуляции родового процесса и лактогенеза. В инициации родового акта фетальному кортизолу, по-видимому, принадлежит ведущая роль. Секрецию глюкокортикоидов контролируют кортиколиберин и адренокортикотропный гормон, или АКТГ. Кортиколиберин — пептидный гормон гипоталамуса, стимулирует секрецию полипептидного гормона аденогипофиза АКТГ, характеризующегося кортикотропной активностью. Простагландин Ф2a (ПгФ2a) — биологически активное соединение с гормональными свойствами, производное полиненасыщенных жирных кислот. Вырабатывается многими клетками и тканями и принимает участие в разнообразных процессах и реакциях организма. У многих домашних животных ПгФ2a служит маточным лютеолитическим фактором (вызывает регрессию желтого тела полового цикла и беременности) и принимает активное участие в инициации и поддержании родового процесса. Желтые тела полового цикла и беременности у собак и кошек мало чувствительны к нему. Аборт у животных данных видов обычно удается вызвать только после многократного применения ПгФ2a, начиная с 5-го дня диэструса. На нейроэндокринную регуляцию половой функции собак существенное влияние оказывает общение с противоположным полом (особенно большое значение это имеет для рефлекторно овулирующих животных — кроликов, верблюдов, лам, представителей семейства кошачьих). Координацию полового поведения самок и самцов обеспечивают органы чувств, кожная чувствительность и движение. Важным внешним фактором, влияющим на все функции организма собак, служит корм. Недостаточное, избыточное и (или) неполноценное кормление не только снижает воспроизводительную способность животных, но часто бывает причиной их бесплодия. Важнейшие репродуктивные функции гормонов гипоталамуса, гипофиза, гонад, плаценты и надпочечников указаны в таблице 2. 1.5. ПОЛОВАЯ И ФИЗИОЛОГИЧЕСКАЯ ЗРЕЛОСТЬПоловая зрелость — это возраст, начиная с которого самцы и самки способны участвовать в процессе полового размножения: продуцировать соответствующие половые клетки (спермии, ооциты) и спариваться. Половая активность у самок носит циклический характер. Готовность к спариванию они демонстрируют только в строго фиксированный период полового цикла — во время половой охоты. Овуляция у собак — спонтанная и привязана к началу половой охоты. Выделение спермы (спермиев и плазмы спермы) из половых органов самцов и введение ее в половые органы самок происходят во время полового акта. Сроки наступления зрелости у самок определяют по дате проявления первого полового цикла. Обычно первый половой цикл регистрируют в возрасте 6…12 мес. У самцов половая зрелость наступает примерно на 1…1,5 мес позднее, чем у самок. Собаки средних и карликовых пород становятся половозрелыми раньше, чем собаки крупных пород. На сроки наступления половой зрелости существенное влияние оказывают состояние здоровья животного, условия кормления и содержания, общение с противоположным полом. Половая зрелость наступает обычно раньше, чем заканчиваются основной рост, структурное и физиологическое развитие животного, обеспечивающие его высокую плодовитость, нормальное функционирование организма в период беременности, родов и лактации, рождение здорового приплода. Физиологическая зрелость — возраст, начиная с которого самцов и самок целесообразно использовать для получения приплода. Физиологическая зрелость у собак по времени совпадает, как правило, с достижением размеров тела, характерных для взрослого животного, и проявлением у самок 2…3 половых циклов. Самки большинства пород становятся физиологически зрелыми обычно в возрасте 1,5 лет, самцы — 2 лет. 1.6. ПОЛОВОЙ ЦИКЛПо ритму полового цикла животных общепринято подразделять на моно- (с одним половым циклом в год) и полициклических, у которых несколько циклов повторяются с определенной для каждого вида животного периодичностью на протяжении всего года или определенного сезона года. Собаки занимают как бы промежуточное положение: на протяжении года у них регистрируют от одного до двух и даже трех половых циклов. По этой причине одни авторы относят собак к моно-, другие — к дициклическим животным. По многим признакам собак следует определять как моноциклических животных. Во-первых, дикие представители семейства собачьих (волки, койоты, шакалы, дикие собаки Динго), с которыми домашняя собака имеет много общих черт и легко скрещивается, — это типичные моноциклические животные, единственный половой цикл которых проявляется в строго фиксированный сезон года. Во-вторых, в половом цикле собак есть характерная для моноциклических животных особенность: течку наблюдают после хорошо выраженного анэстрального периода. В процессе одомашнивания половая цикличность собак изменилась: теперь у них регистрируют несколько половых циклов в течение года и вне зависимости от сезона (Sokolowski J.H., 1977). Исключение составляют собаки породы басенджи, единственный половой цикл которых проявляется обычно осенью. Указанная особенность, по-видимому, обусловлена действием простого рецессивного гена, поскольку при скрещивании собак породы басенджи с собаками других пород у одних потомков отмечают один половой цикл в год, у других — несколько (McDonald L.E., 1980). В повседневной практике половую активность самок учитывают по межэстральному периоду, так как определить продолжительность полового цикла и, в частности, время его окончания можно только с помощью лабораторных методов исследований: по динамике прогестерона в крови и картине влагалищных мазков. Продолжительность межэстрального периода сильно варьирует и зависит от породы собаки, но не от ее размеров (табл. 3). 3. Продолжительность межэстрального периода у собак некоторых пород (Sokolowski J.H., 1977) Беременность не оказывает существенного влияния на продолжительность межэстрального периода: у самок породы бигль его продолжительность при наступлении беременности в среднем составляет 245,1 дня, при ее отсутствии — 235,8 дня (Sokolowski J.H., 1977). Половой цикл собак уникален, он по многим признакам отличается от цикла сельскохозяйственных и лабораторных животных. Наступлению половой охоты у собак предшествуют геморрагические выделения, которые возникают в результате диапедеза (просачивания клеток крови через стенку кровеносного сосуда). Яйцеклетка овулирует в начале половой охоты незрелой (на стадии ооцита первого порядка) и способность к оплодотворению приобретает через 2…3 дня после попадания в яйцевод. Функционально активное желтое тело образуется к моменту окончания половой охоты. У большинства животных других видов желтое тело формируется в конце половой охоты или после ее окончания, и этот период принято выделять в самостоятельную стадию полового цикла (метэструс, или стадию торможения). После регрессии желтого тела полового цикла и беременности наступает анэструс (период полового покоя). Видимо, логично делить межэстральный период собак на следующие четыре стадии: проэструс, эструс, диэструс и анэструс. Стадию метэструса у собак следует рассматривать как постовуля-ционную фазу эструса. Бессмысленно ее выделять в самостоятельную стадию, поскольку период овуляции и образования желтого тела у собак не удается диагностировать клиническими методами и он представляет для исследователей только академический интерес. Благодаря делению межэстрального периода на проэструс, эструс, диэструс и анэструс можно избежать путаницы в научной терминологии и обозначить соответствующие стадии полового цикла у животных разных видов одними и теми же терминами. Обзор терминов и критериев начала и окончания стадий полового цикла и анэструса приведен в таблице 4. 4. Терминология и критерии начала и окончания стадий полового цикла и периода полового покоя у собак по данным разных авторов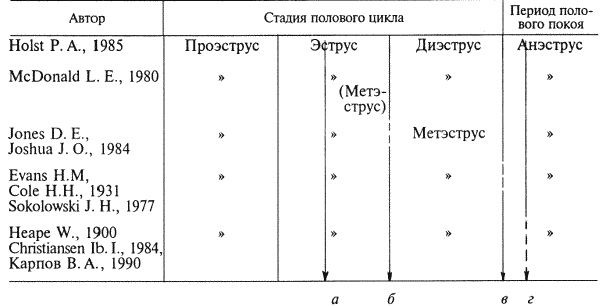 Примечание: а — время овуляции; б — время окончания половой охоты; в — время окончания функционирования желтого тела полового цикла; г — время завершения регенерации эндометрия. Проэструс — стадия подготовки половых органов и всего организма самки к половому акту, продолжительность которой составляет в среднем 9 (2…16) дней. Проэструс у собак хорошо выражен, его легко диагностировать по признакам течки и полового возбуждения. ФСГ стимулирует развитие преовуляторных фолликулов. Про-гестагенная активность яичников находится на самом низком уровне. Концентрация прогестерона в плазме периферической крови ниже 1?10-9 г/мл. Эстрогенная активность яичников по мере развития преовуляторных фолликулов возрастает. Максимальную концентрацию эстрогенов (преовуляторный пик 17?-эстрадиола) в плазме периферической крови регистрируют в конце проэструса или в начале эструса. Эстрогенные гормоны, воздействуя на ЦНС и половые органы, обусловливают развитие признаков течки и полового возбуждения. Течка — процесс выделения слизи из половых органов самки. У собак течка характеризуется следующими признаками: половая петля (вульва) увеличена в размере, набухшая; слизистые оболочки влагалища и его преддверия гиперемированы и отечны; сократительная активность матки повышена. Все слои стенки матки утолщены и обильно кровоснабжены. Шейка матки расслаблена, цервикальный канал раскрыт. Чрезмерная васкуляризация эндометрия приводит к диапедезу клеток крови в просвет матки и появлению геморрагических выделений, которые к концу проэструса становятся более светлыми и менее обильными. Существенные изменения претерпевает эпителий влагалища и его преддверия. Толщина эпителиального слоя резко возрастает, поверхностные эпителиальные клетки ороговевают. Ороговевшие клетки выполняют защитную функцию. Они очень устойчивы к бактериальной инвазии, действию химических и физических факторов. В мазках, взятых из краниальной части влагалища в период проэструса, обнаруживают вариабельное количество лейкоцитов, много эритроцитов и ороговевших эпителиальных клеток без ядра или с пикнотическим ядром. Половое возбуждение — изменение в поведении животного. В период проэструса у самок появляется жажда, извращается чутье, они становятся беспокойными, раздражительными, плохо выполняют команды, могут убежать от владельца; обычно часто и в разных местах мочатся — как бы метят территорию, выделяя с мочой ароматические (пахучие) вещества — половые феромоны, привлекающие самцов. Самцы проявляют повышенный «интерес» к самкам с признаками течки и полового возбуждения и беспрерывно преследуют их. «Интерес» самок к самцам обычно возникает только в конце проэструса: самки становятся игривыми, но не допускают садки самца и полового акта. Эструс — период половой охоты: положительной сексуальной реакции самки на самца. Половая охота у собак длится в среднем 9 (2…16) дней. Выделения из половой щели, как правило, светлые или со следами крови. Половые губы становятся менее отечными и более ригидными. Самка проявляет повышенный «интерес» к самцам, обнюхивает их половые органы; затем выбирает самца-фаворита, допускает садку и коитус. В начале эструса у самок происходит циклический выброс ЛГ, инициируемый преовуляторным пиком 17? — эстрадиола. Яйцеклетка обычно овулирует на 2…5-й день половой охоты: примерно через 30 ч после того как концентрация ЛГ достигнет максимального значения. На месте овулировавших фолликулов образуются желтые тела, которые становятся функционально активными к концу эструса. После овуляции прогестагенная активность яичников возрастает, а эстрогенная, наоборот, снижается. Во влагалищных мазках в начале эструса отмечают единичные эритроциты и лейкоциты и резкое преобладание ороговевших эпителиальных клеток над эпителиальными клетками, содержащими интактное ядро. Наиболее высокий индекс ороговения во влагалищных мазках регистрируют в период максимальных концентраций ЛГ (преовуляторный пик ЛГ). Диэструс — период функционирования желтого тела полового цикла. Начинается с момента окончания половой охоты и заканчивается регрессией желтого тела. Продолжительность диэструса может сильно варьировать (от 40 до 84 дней) и в среднем составляет 60 дней. После окончания половой охоты у самок достаточно быстро исчезают признаки течки и полового возбуждения. Животные становятся спокойными. Во влагалищных мазках преобладают промежуточные и парабазальные эпителиальные клетки, содержащие ядро. Диэструс — самая активная часть лютеиновой фазы полового цикла, начинающаяся сразу же после овуляции. Концентрация прогестерона в плазме периферической крови увеличивается и на 30-й день после проявления преовуляторного пика ЛГ достигает максимальных значений — 25…30?10-9 г/мл (Concannon P. W., 1986). Затем содержание прогестерона начинает медленно снижаться. Механизм регрессии желтого тела полового цикла у собак не выяснен. Анэструс — период полового покоя. Его продолжительность может сильно варьировать и в среднем составляет 125 дней. В этот период яичники мало активны. Рост фолликулов носит абортивный (незавершенный) характер. Третичные фолликулы на разных стадиях своего развития подвергаются атрезии. Идет активный лизис желтых тел предыдущего полового цикла. В эндометрии восстанавливается (регенерирует) эпителиальный слой. Во влагалищных мазках встречаются только парабазальные и промежуточные эпителиальные клетки. 1.7. ПОЛОВОЙ АКТПоловой акт, или совокупление, коитус, копуляция, — совокупность половых рефлексов, обеспечивающих выведение спермы из полового аппарата самца и введение ее в половые пути самки. В половом акте различают следующие рефлексы: приближения, эрекции, обнимательный, совокупительный и эякуляции. Рефлекс приближения — совокупность поведенческих реакций, координирующих сексуальное поведение разнополых особей при спаривании. Рефлекс эрекции — сильное наполнение кровью и увеличение полового члена в размере, обеспечивающие его выведение из препуция и возможность введения в половые пути самки. Этот рефлекторно-сосудистый акт проявляется у самок набуханием клитора и гиперемией слизистой оболочки влагалища и его преддверия. Обнимательный рефлекс — принятие самкой и самцом позы для спаривания. Одновременно с обнимательным проявляется и совокупительный рефлекс. Совокупительный рефлекс — введение и фрикция полового члена самца во влагалище самки, приводящие к раздражению эрогенных зон их гениталий и наступлению эякуляции. Рефлекс эякуляции — выведение спермы (эякулята) из полового аппарата самца. Эякуляция, так же как и эрекция, это нейрорефлекторный акт. Все половые рефлексы по своей природе врожденные (безусловные) В процессе их становления и приобретения животными сексуального опыта на безусловные рефлексы наслаиваются условные. Одни из них способствуют развитию полноценных половых рефлексов, другие, наоборот, усиливая или ослабляя их проявление, приводят к возникновению анормального полового поведения. Сексуальное поведение животных координируют органы чувств кожная чувствительность и движение. В период проэструса и эструса самки выделяют половые феромоны и в частности, метил-п-гидроксибензоат, стимулирующие половые рефлексы самцов. В начале проэструса самки ведут себя пассивно или же демонстрируют по отношению к самцам агрессивное поведение, в конце проэструса начинают проявлять к ним сексуальный интерес. Самцы следуют за самкой с признаками течки и полового возбуждения, выстроившись по иерархическому рангу Если самка позволяет, то самец-фаворит начинает активно обнюхивать ее, облизывать наружные половые органы и другие участки ее тела, кладет голову, одну или две передние лапы ей на спину, предпринимает попытки совершить половой акт. При наступлении половой охоты самка демонстрирует готовность к спариванию. Обнюхивает половые органы самца, принимает позу для спаривания: стоит неподвижно, отводит в сторону хвост, подтягивает кверху половую петлю. Самец вспрыгивает на самку, обхватывает передними лапами ее за бока и совершает толкательные движения тазом, обеспечивающие введение полового члена во влагалище (рис. 11). 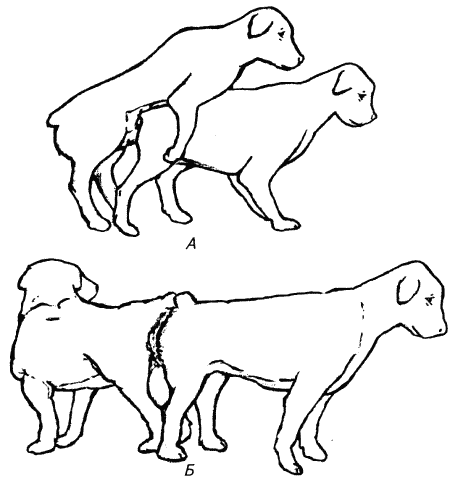 Рис. 11. Позы собак при спаривании: А — в начале полового акта; Б — в период сцепления половых органов При полном введении пениса рефлекторно сжимается сфинктер преддверия влагалища, что приводит к сильному набуханию луковицы полового члена и сцеплению (склещиванию) половых органов животных. Самец спрыгивает с самки, и животные принимают позу «хвост к хвосту». Сцепление половых органов (половой замок, вязка) продолжается 5…45 мин. Выделение спермы сопровождается ритмичными движениями корня хвоста и волнообразными сокращениями стенки мочеполового канала. Сперма выделяется во влагалище в виде трех фракций: первая фракция выполняет функцию смазки, вторая содержит спермии, третья обеспечивает проталкивание спермиев в полость матки. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||