|
||||
|
|
ГЛАВА 1 Особенности физиологии и эндокринологии сук И. Джеффкоут (Ian Jeffcoate) ОБЩИЕ СВЕДЕНИЯ ОБ ЭСТРАЛЬНОМ ЦИКЛЕ Эстральный цикл представляет собой последовательность взаимосвязанных изменений, происходящих как в яичниках, матке, влагалище, так и в поведении животного. Подобные изменения, характерные для всех млекопитающих, обеспечивают созревание яйцеклеток и внутриматочное развитие эмбрионов. У большинства небеременных самок домашних животных эстральный цикл включает следующие стадии: эструс — период, когда созревшие яйцеклетки находятся в маточных трубах, а половая рецептивность повышает вероятность оплодотворения; диэструс — период подготовки организма к беременности. Если оплодотворения не произошло, наблюдается возврат к проэструсу, во время которого возобновляется развитие фолликулов, приводящее к новому эструсу и соответственно к половой охоте. У некоторых животных — свиней, крупного рогатого скота и лошадей — эструс возобновляется в среднем каждые 3 недели, но в случае беременности наблюдается достаточно продолжительный диэструс, необходимый для ее развития. Цикл домашней собаки во многом отличается от этой общей схемы. Во-первых, его продолжительность составляет не менее 5 месяцев; во-вторых, беременность развивается в период нормального диэструса, но не продлевает его; в-третьих, между циклами независимо от наличия или отсутствия беременности наступает так называемый анэструс, т. е. продолжительный период относительного покоя яичников. Указанные особенности эстрального цикла суки отражены в специфической терминологии, требующей некоторых пояснений. Поскольку у большинства видов проэструс очень короткий, его длительностью, как правило, пренебрегают. Термин «диэструс» (буквально «между эструсами») часто, хотя и некорректно используют для обозначения межэстрального интервала у собак. Однако если у других млекопитающих этот интервал включает лютеиновую фазу и фазу проэструса, то у сук он включает не только лютеиновую фазу, но также и весьма продолжительный анэструс; исходя из этого лютеиновую фазу предпочтительнее обозначать как «метэструс», а не «диэструс». Несмотря на указанные различия, эстральный цикл у сук регулируется теми же эндокринными процессами, что и у других видов животных. • Основной контроль обеспечивают гипоталамус и гипофиз; • Яичники вырабатывают стероидные гормоны — эстрогены (главным образом эстрадиол) и прогестерон; • Половые гормоны воздействуют на репродуктивный тракт, добавочные половые железы и участки мозга, управляющие репродуктивным поведением. Настоящая глава знакомит с общими закономерностями репродуктивного цикла у сук и описывает ситуации, в которых целесообразно проведение эндокринологических исследований и других активных вмешательств. ПРОЭСТРУС О наступлении проэструса свидетельствуют специфические симптомы, к которым относятся покраснение и набухание вульвы с последующим появлением слизисто-кровянистых выделений. Принято считать, что эти выделения появляются в первый день проэструса. Одновременно наблюдаются такие поведенческие изменения, как учащение мочеиспускания, беспокойство и непослушание, склонность к бродяжничеству; отмечается также усиление реакции самца на самку. Эти перемены объясняются повышением концентрации эстрадиола в сыворотке крови (фиг. 1.1), связанным с активизацией фолликулов яичников. Эстрадиол запускает целый ряд процессов в репродуктивном тракте, включая рост железистого эпителия, васкуляризацию слизистых и их отек. Наличие кровянистых выделений объясняется повышением хрупкости капилляров в отечной слизистой и поступлением крови в полость матки. Отек половых путей, характерный для периода проэструса, визуализируется на ультразвуковом изображении как их общее сужение. Кроме того, разрастание слизистой приводит к покраснению и вздутию ее складок, что ясно видно при эндоскопии. Под действием эстрадиола происходит также активная пролиферация клеток эпителия влагалища. Морфологические изменения влагалища будут рассмотрены ниже в связи с методами определения момента овуляции и оптимальных сроков оплодотворения. 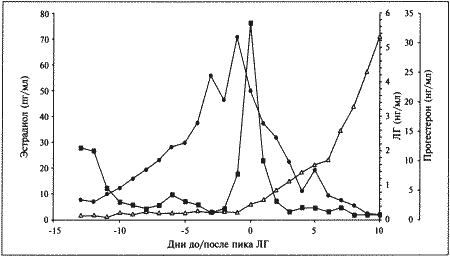 Фиг. 1.1. Фиг. 1.1. Изменения концентрации эстрадиола (?), лютеинизирующего гормона (ЛГ,) и прогестерона (?) в сыворотке крови у сук. На графике за точку отсчета принят день пика концентрации ЛГ. Следует отметить, что пик концентрации эстрадиола приходится на день минус 1, уже на день 0 заметно повышение концентрации прогестерона. (Перевод единиц: 1 пг/мл эстрадиола = 3,6 пмоль/л; 1 нг/мл прогестерона = 3 нмоль/л) Стадия проэструса характеризуется развитием фолликулов под влиянием гонадотропных гормонов — лютеинизирующего (ЛГ) и фолликулостимулирующего (ФСГ), выделяемых передней долей гипофиза. Секреция этих гормонов регулируется гипоталамическим гонадотропинрилизинг гормоном (ГнРГ). У собак, так же, как и у других животных, вслед за пиком концентрации эстрадиола возникает (спустя сутки или чуть больше) пик концентрации ЛГ (фиг. 1.1). Таким образом, выброс ЛГ происходит всегда автоматически вслед за повышением активности фолликулов, вырабатывающих эстрадиол, и поскольку овуляция следует за возникающим без внешней стимуляции пиком концентрации ЛГ, ее называют спонтанной. Описанная ситуация разительно отличается от картины, которую можно наблюдать у кошек, для которых характерна индуцированная овуляция: у них выброс ЛГ и овуляция происходят вслед за коитусом. Насколько можно судить, у таких животных эстрадиол не способствует активации секреции ЛГ. Повышение уровня ФСГ необходимо для стимуляции развития фолликулов и секреции эстрадиола, однако концентрация ФСГ в крови не достигает столь высоких значений, какие наблюдаются при пике ЛГ; это объясняют действием ингибина — белка, секретируемого фолликулами, который избирательно подавляет секрецию ФСГ Тем не менее ФСГ играет важную роль в созревании фолликулов и подготовке их к трансформации в желтые тела после овуляции. Начало трансформации фолликулярных клеток рассматривают как ключевой момент всего цикла, поскольку оно сопровождается преовуляторным повышением секреции прогестерона (фиг. 1.1), которое, по-видимому, играет ведущую роль в пусковом механизме овуляции и подтверждении эструса. Наблюдаемый у собак преовуляторный рост концентрации прогестерона нетипичен для других домашних животных, в частности для жвачных, у которых наступление эструса и овуляции происходит на фоне минимального уровня прогестерона. Однако преовуляторная концентрация прогестерона у собак все же относительно низка, и следует помнить, что назначение высоких доз прогестагенов в период проэструса вызывает отсрочку эструса и овуляции. Овуляторный пик ЛГ обычно описывают как кульминацию цикла, что объясняется его ролью в процессах овуляции и перехода к метэструсу, т. е. к лютеиновой фазе, поддерживаемой прогестероном. По причинам, изложенным выше, а также потому, что момент наступления эструса у разных сук варьирует, для оптимального подсчета сроков беременности за точку отсчета принимают пик ЛГ, а не первый день эструса, как практикуется в работе с другими видами животных. ЭСТРУС Половые гормоны и поведение Термин «эструс» образован от латинского «oestridae», что переводится как «овод» и буквально означает «безумное поведение», имеется в виду, что если в период проэструса сука привлекательна для кобелей, но не подпускает их к себе, то с переходом к стадии эструса она демонстрирует половую охоту, принимая характерные позы (лордоз), свидетельствующие о готовности к спариванию. Под воздействием эстрадиола в организме суки вырабатываются специфические сигнальные вещества — феромоны, воспринимаемые кобелем посредством органов обоняния. Феромоны вырабатываются почками и половым трактом, они выводятся из организма с мочой и вагинальными выделениями и специально предназначены для обозначения сексуального статуса. В сочетании с поведенческими проявлениями они усиливают сексуальную привлекательность суки и стимулируют репродуктивную активность самцов. Одним из феромонов является метиловый эфир 4-гидроксибензойной кислоты; обработка им половых органов суки в период анэструса или даже кастрированной суки стимулирует возбуждение и попытки спаривания у кобелей. Эффект феромонов распространяется не только на кобелей, но и на других сук, у которых они могут вызвать ускоренное наступление эструса и синхронизировать половой цикл, в особенности при групповом содержании собак. Данный факт свидетельствует о том, что феромоны способны влиять на гипоталамическую секрецию ГнРГ и тем самым в конечном итоге стимулировать активность яичников. Предполагается, что высокие концентрации эстрадиола запускают активность участков мозга, отвечающих за репродуктивное поведение, однако для полного развития нормального для эструса поведения требуется прекращение эстрогенного действия на фоне присутствия прогестерона. Ановуляторные циклы (при которых фолликулы не овулируют и не вступают в лютеиновую фазу с ее высокой секрецией прогестерона) характеризуются поведением, типичным для проэструса, которое постепенно затухает, но вновь возвращается позже, если, минуя лютеиновую фазу, развивается новая когорта фолликулов, продуцирующих эстрогены. Подобное нарушение называют расщепленной течкой. Овуляция Повышение концентрации прогестерона является необходимым условием овуляции; соответственно начальное увеличение концентрации этого гормона в плазме происходит непосредственно перед овуляцией и может использоваться как индикатор ее приближения. Как правило, у сук наблюдается множественная овуляция; согласно данным гистологических и лапароскопических исследований, в большинстве случаев овуляция происходит в течение 30–48 часов после пика концентрации ЛГ, хотя часть фолликулов овулируют лишь через 96 часов. После овуляции, но до оплодотворения ооциты проходят очередную стадию мейоза. Этот процесс с трудом поддается изучению, однако тщательный хронометраж искусственного осеменения замороженной спермой, взятой от разных кобелей (для упрощения идентификации потомства), приводит к выводу, что созревание ооцитов занимает 2–3 дня. Если сложить их с количеством дней, отделяющих пик концентрации ЛГ от овуляции, то получится, что оплодотворение возможно не раньше, чем через 4 дня, считая от пика концентрации ЛГ; этот день является началом фертильного периода (фиг. 1.2). Ошибки в его вычислении объясняются тем, что признаки эструса наблюдаются уже за 5 дней до начала фертильного периода и продолжаются еще 5 дней после его окончания. Следующие факторы определяют длительность фертильного периода: длительное (до нескольких дней) сохранение жизнеспособности спермы в половом тракте суки; время, необходимое для овуляции и созревания ооцитов (2–3 дня) при сохранении жизнеспособности яйцеклеток (1–2 дня). С учетом этих факторов фертильным следует признать период, продолжающийся от 4-го до 7-го дня, считая от пика концентрации ЛГ. При стандартном подсчете продолжительность беременности составляет от 58 до 72 дней, но если принять за точку отсчета овуляторный пик ЛГ, длительность беременности составит 65 дней с очень небольшими отклонениями. Оплодотворение Первичное воздействие эстрогенов на слизистую оболочку стимулирует секреторную активность слизистой, а это способствует выживанию и транспортировке гамет. Известно также, что эстрогены повышают возбудимость гладкой мускулатуры, но, вероятно, не прямо, а через адренергические рецепторы. Это влияние эстрогенов совместно с активностью реснитчатого эпителия половых путей обеспечивает продвижение сперматозоидов и доставку созревших ооцитов в маточные трубы. Позднее, на этапе лютеинизации фолликулов возрастающая концентрация прогестерона приводит к такому изменению характера секреции слизистой, которое препятствует дальнейшему продвижению и выживанию сперматозоидов в половом тракте самки (в частности, их миграцию через шейку матки) (см. ниже). Кроме того, прогестерон уменьшает возбуждение гладкой мускулатуры и тем самым замедляет прохождение яйцеклеток или эмбрионов в маточных трубах, задерживая эмбрионы до тех пор, пока в матке не создадутся благоприятные условия для их имплантации. 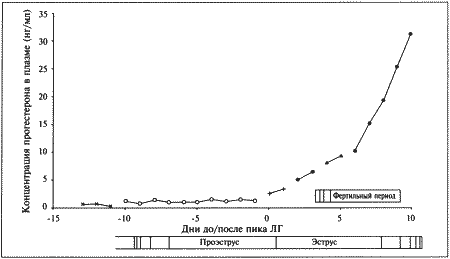 Фиг. 1.2. Фиг. 1.2. Динамика концентрации прогестерона в плазме крови на протяжении эстрального цикла. Составлено на основании данных, полученных при наблюдении за 8 собаками. За точку отсчета принят овуляторный пик ЛГ. Ключевые показатели, используемые для идентификации стадии цикла и вычисления фертильного периода (см. текст): концентрация прогестерона в конце анэструса находится на базальном уровне (может не выявляться) (х-х), затем в период проэструса незначительно повышается до определяемого уровня (о-о); в период, предшествующий овуляции, концентрация прогестерона возрастает одновременно с пиком ЛГ (+-+). За временной интервал между пиком ЛГ (^-^) и овуляцией уровень прогестерона повышается до характерных значений начала фертильного периода и продолжает увеличиваться (-•) приблизительно до 30 дня. Табл. 1.1. Время проявления основных клинических признаков (по отношению к пику ЛГ) по данным наблюдения за 8 собаками.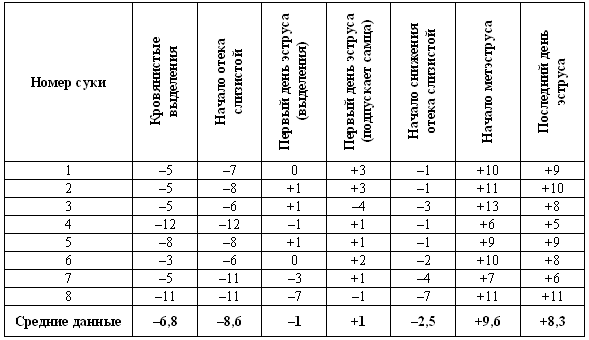 Предполагаемый фертильный период — от 4-го до 7-го дня. Примечания: — кровянистые выделения появляются за несколько дней до начала эструса; — самка подпускает к себе самца непосредственно перед началом фертильного периода; — последний день восприимчивости может наблюдаться спустя несколько дней после окончания фертильного периода, т. е. в начале метэструса; — начало уменьшения отека слизистой оболочки обычно наблюдается перед пиком ЛГ. Эти хорошо известные функции двух главных стероидных гормонов, вырабатываемых яичниками, имеют равное значение для сук, несмотря на сравнительно более раннее изменение соотношения эстроген/прогестерон в результате преовуляторной лютеинизации (фиг. 1.1). На основании вышесказанного мы можем заключить, что в организме сук прогестерон обеспечивает выживание, транспорт и оплодотворение гамет, однако все это требует предварительного повышения концентрации эстрадиола (проэструс). У собак — как и у других видов — изменение соотношения эстроген/прогестерон в результате приема после овуляции фармакологических препаратов на основе эстрогенов увеличивает проходимость маточных труб, что приводит к слишком раннему попаданию эмбрионов в матку. На этом явлении основана посткоитальная эстрогеновая терапия. ВЫЧИСЛЕНИЕ ФЕРТИЛЬНОГО ПЕРИОДА Учитывая долгий период сексуальной рецептивности у сук, включающий овуляцию и период созревания гамет, фертильным следует считать интервал между 4 и 7 днем после пика концентрации ЛГ. Однако не исключено, что ближе к окончанию фертильного периода способность к оплодотворению снижается, тогда как некоторые ситуации требуют особенно надежного вычисления (например, искусственное осеменение замороженной спермой). Для определения фертильного периода и оптимальных сроков вязки применяют различные методы, к которым относятся вагинальная цитология, вагинальная эндоскопия и анализ концентрации гормонов в сыворотке крови. Вагинальная цитология Повышение концентрации эстрадиола в период проэструса стимулирует деление клеток в базальных слоях вагинального эпителия, но затем концентрация эстрадиола, а соответственно и эндокринная поддержка образования нового, многослойного эпителия снижается, поэтому в образце обнаруживается больше мертвых ороговевших клеток. По мере развития проэструса снижается количество эпителиальных клеток, содержащих ядро. С начала до середины проэструса в вагинальных мазках обнаруживают повышенное содержание эритроцитов. Пик ороговения совпадает с началом повышения концентрации прогестерона; однако в начале эструса исследование не обнаруживает характерных особенностей, которые могли бы указывать на начало фертильного периода. Как правило, такой период наступает несколькими днями позже (табл. 1.1). Тем не менее вагинальная цитология остается популярным методом определения репродуктивного статуса суки из-за своей дешевизны, простоты и доступности. Для получения образца используют ватный тампон, предпочтительно небольшой, который вводят во влагалище и осторожно берут мазок. Полученные клетки помещают под микроскоп, а затем окрашивают контрастным (трихромом) или неконтрастным (например, препаратом Diff-Quik) веществом. При таком окрашивании мертвые кератинизированные клетки становятся оранжевыми, тогда как активные ядерные клетки, а также базальные и парабазальные эпителиальные клетки приобретают различные оттенки от голубого до зеленого. Ближе к окончанию эструса вагинальные выделения претерпевают характерные изменения (снова выявляются эпителиальные клетки, содержащие ядро, и появляется большое количество лейкоцитов). Такая картина обычно наблюдается через 7–9 дней после пика ЛГ и известна как «вагинальный мазок метэструса». Изменению характера выделений предшествует переходный период, характеризующийся возрастающим количеством активных клеток и указывающий на окончание фертильного периода и эструса (табл. 1.1). Вагинальная цитология весьма информативна в плане определения фазы эстрального цикла, однако значительные расхождения во времени появления основных признаков эструса по отношению к пику фертильности ограничивает применение указанной методики. Вагинальная эндоскопия Прекращение секреции эстрадиола фолликулами и переход их к секреции прогестерона приводят к снижению как отечности, так и васкуляризации слизистых оболочек, что сопровождается выраженным изменением характера вагинальных выделений, выявляемым с помощью вагинальной эндоскопии. На стадии проэструса слизистая оболочка становится более выпуклой и отечной, затем наблюдается ее уменьшение; по мере приближения овуляторного пика слизистая сжимается и бледнеет, в некоторых случаях становясь практически белой (фиг. 1.1). Перечисленные изменения свидетельствуют о приближении стадии, характеризующейся постепенным повышением концентрации прогестерона перед овуляцией и последующим наступлением фертильного периода. Эти изменения легко узнать, имея даже небольшую практику (фиг. 1.3), кроме того, они настолько воспроизводимы, что по набору признаков их можно оценивать полуколичественно. Вагинальная эндоскопия весьма полезна в определении оптимальных сроков вязки. По окончании фертильного периода, т. е. в начале метэструса — слизистая оболочка влагалища бледнеет и истончается, складки становятся закругленными — и что, вероятно, является наиболее характерным признаком, — слизистая в переднем отделе влагалища выглядит раздраженной и при прикосновении быстро сжимается, образуя розетку. Измерение концентрации гормонов Данные о концентрации прогестерона в сыворотке крови могут служить ориентиром для определения фертильного периода у сук. Механизмы и динамика изменения концентрации прогестерона описаны в предыдущем разделе (фиг. 1.2). В конце анэструса концентрация прогестерона в сыворотке крови находится на базальном уровне (и практически не определяется), к окончанию проэструса она повышается до низкой, но поддающейся определению отметки (около 3 нмоль/л или 1 нг/мл). И продолжает повышаться, достигая уровня 3–6 нмоль/л (1–2 нг/мл) к моменту овуляторного пика ЛГ; 2 дня спустя (в день овуляции) показатели достигают 6–12 нмоль/л (2–4 нг/мл), а к 4 дню (т. е. к началу фертильного периода) — 18–30 нмоль/л (6–10 нг/мл) (фиг. 1.1). Концентрацию прогестерона определяют в плазме крови, поскольку анализ других жидкостей (слюны и мочи) в настоящее время не дает точных результатов. Исследование проводят после появления первых признаков проэструса и повторяют по меньшей мере каждые 2–3 дня до окончания проэструса или начала поведенческого эструса. Такая методика позволяет выявлять стремительное нарастание концентрации прогестерона перед овуляцией и соответственно рассчитать дату возможной вязки. В случаях очень короткого проэструса исследование начинают раньше. В зависимости от метода исследования (радиоиммунный или иммуноферментный) возможны некоторые расхождения в показателях концентрации прогестерона; при этом следует учитывать, что иммуноферментный метод дает завышенные, но в остальном надежные результаты. Большая часть опубликованных данных (включая данные, приведенные в настоящей главе) получены с помощью радиоиммунного метода. Готовые иммуноферментные тест-системы, предназначенные для определения концентрации прогестерона в плазме крови на различных стадиях эстрального цикла, дают результаты, почти вдвое превосходящие данные радиоимунного анализа (РИА), но в той же степени отражающие характерные циклические изменения. С помощью готовых наборов нельзя получить абсолютные показатели концентрации прогестерона, но можно проследить относительные изменения его концентрации от начала проэструса и до конца цикла. 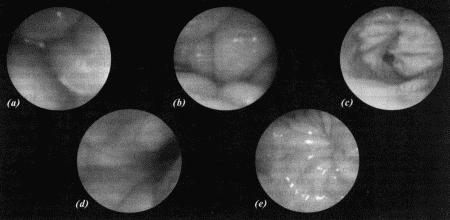 Фиг. 1.3. Фиг. 1.3. Эндоскопическое исследование слизистой влагалища. Динамика изменений на протяжении цикла: (а) проэтрус — розовая окраска и отечность; (b) начало эструса — слизистая бледнеет, отек начинает уменьшаться (обычно перед пиком ЛГ); (c) середина эструса — слизистая бледная, уменьшение отека (сморщивание) явно выражено, что соответствует середине фертильного периода; (d) начало метэструса — видны закругленные складки, при прикосновении слизистая смыкается, образуя розетку (е) (см. Приложение) Прогестерон является единственным гормоном, концентрацию которого используют как ориентир при вычислении фертильного периода. Хотя ЛГ и эстрадиол в этом отношении даже более информативны, их определение не практикуется. Другие аспекты определения фертильного периода Последние рентгенографические исследования влагалища с применением контрастного вещества выявили устойчивую связь между концентрациями эстрадиола и прогестерона в крови и раскрытием шейки матки. Результаты этих исследований имеют решающее значение для определения фертильного периода у собак, поскольку шейка матки остается закрытой (непроницаемой для контрастного вещества и, вероятно, сперматозоидов) до тех пор, пока не произойдет снижение концентрации эстрадиола и не вырастет соотношение прогестерон/эстрадиол во время преовуляторного периода. После пика ЛГ шейка матки остается открытой в течение приблизительно 6 дней (т. е. она закрывается примерно за 2 дня до начала цитологического метэструса), и в этот период концентрация прогестерона остается высокой, а эстрадиол практически не определяется. Приведенные данные свидетельствуют о том, что сперматозоиды имеют доступ в шейку матки только в ограниченное время эструса вопреки общепринятому мнению, согласно которому вязку можно проводить как до, так и после раскрытия шейки матки. Изменения состояния шейки матки объясняются утолщением слизистой оболочки под воздействием эстрогена на стадии проэструса, что приводит к закрытию шейки матки. Последующее раскрытие шейки матки сопровождается снижением отечности слизистой оболочки и обусловлено увеличением соотношения прогестерон/эстрадиол. Интересно, что период, когда шейка матки раскрыта, соответствует оптимальным срокам вязки, рассчитанным на основании результатов эндоскопического исследования слизистой влагалища. Данные рентгенографических исследований с применением контрастного вещества удобны для интерпретации, однако не объясняют, каким образом на стадии проэструса кровь и маточный секрет проникают через отечную шейку матки. Вероятно, сказывается влияние других активаторов, например, эстрогенов, способных расслаблять гладкую мускулатуру, а также простагландинов, содержащихся в сперме. Необходимость повышения концентрации прогестерона для наступления эструса и индукции овуляторного пика ЛГ указывает на то, что устойчивые проявления течки наблюдаются лишь непосредственно перед началом фертильного периода. Хотя начало течки по отношению к пику ЛГ — индивидуальная характеристика особи (табл. 1.1), указанные проявления могут служить ориентиром для вычисления фертильного периода. МЕТЭСТРУС Фаза метэструса следует за эструсом; ее начало определяют по снижению полового возбуждения у самки (обычно на 6–8 день после начала эструса или 8–10 день после преовуляторного пика ЛГ). Переход к метэструсу сопровождается изменением характера вагинальных выделений (повышается содержание лейкоцитов и возрастает количество клеток, содержащих ядра); эндоскопия выявляет сокращение и истончение профиля слизистой влагалища, свидетельствующие об окончании фертильного периода. Специфических изменений эндокринного статуса не наблюдается, за исключением неуклонного, продолжающегося от начала эструса, повышения концентрации прогестерона до 30–90 нмоль/л (10–30 нг/мл) (фиг. 1.2). Как указано выше, беременность развивается в фазе метэструса, т. е. в условиях доминирования прогестерона. Как при наличии, так и в отсутствие беременности прогестерон вырабатывается исключительно желтыми телами; плацента в секреции прогестерона не участвует. На стадии метэструса происходит развитие эмбрионов; интересно отметить согласованность протекания некоторых основных этапов, предшествующих имплантации, и продемонстрировать их связь с изменениями концентрации прогестерона и эстрадиола (фиг. 1.1). Наиболее целесообразно принять за точку отсчета овуляторный пик ЛГ. Как уже упоминалось, овуляция происходит на 2 день, после чего яйцеклетки проходят период созревания. К 4–7 дню они готовы к оплодотворению. Оплодотворенные яйцеклетки некоторое время удерживаются в маточных трубах, они проходят маточно-трубное соединение и приблизительно на 10 день перемещаются в рога матки. Увеличение соотношения прогестерон/эстрадиол (фиг. 1.1), судя по всему, имеет жизненно важное значение для этого этапа. Бластоцисты локализуются в роге матки вплоть до 13 дня, после чего свободно мигрируют из одного рога в другой до 16 дня, когда набухание эндометрия свидетельствует о том, что имплантация эмбрионов произошла. С начала метэструса концентрация прогестерона продолжает нарастать, достигая максимума (порядка 90–270 нмоль/л; 30–90 нг/мл) к 30 дню от пика ЛГ, а затем постепенно снижается до 30 нмоль/л (10 нг/мл) к 60 дню от пика ЛГ. Таким образом, концентрация прогестерона в крови сохраняется на уровне 30–90 нмоль/л (10–30 нг/мл) около 2 месяцев после окончания эструса; более низкий уровень указывает на ановуляторный цикл или дисфункцию желтых тел. Вплоть до 60 дня время и диапазон колебаний концентрации прогестерона у беременных и небеременных самок одинаковы. Различия проявляются в резком снижении концентрации прогестерона перед родами (см. ниже), тогда как у небеременных самок, у которых отсутствует лютеолитический механизм, концентрация прогестерона снижается постепенно и достигает 3–9 нмоль/л (1–3 нг/мл) в течение последующих 30–60 дней. Неизвестно, проявляется ли активность желтых тел в строго запрограммированный отрезок времени или спад активности объясняется снижением лютеотрофической поддержки. Повышение концентрации пролактина в сыворотке, отмечающееся как у беременных, так и у небеременных сук в период с 30 по 65 день после пика ЛГ, сохраняется до 60–90 дня, что особенно характерно для кормящих сук, поскольку пролактин обеспечивает секрецию молока (но не его количество). В некоторых случаях у сук может наблюдаться ложная лактация, развивающаяся спонтанно, или как следствие кастрации, проведенной на стадии метэструса. К вероятным причинам данной патологии следует отнести повышение концентрации пролактина в конце метэструса (в отсутствие беременности) в связи с недостаточным угнетением его выработки из-за повышения концентрации прогестерона. Кроме того, можно предположить, что прогестерон уменьшает чувствительность молочных желез к пролактину, а лактация в конце метэструса обусловливается снижением концентрации прогестерона, вырабатываемого желтым телом. Вероятно, эта особенность объясняется происхождением собак от диких предков: при стайном проживании циклы доминирующих самок синхронизируются с циклами нерожавших самок, которым обычно поручается воспитание щенков. Прерывание беременности воздействием на секрецию прогестеронаЭкспериментальные исследования показывают, что первоначально желтые тела практически автономны и в течение 20 первых дней метэструса способны вырабатывать прогестерон без лютеотропной поддержки со стороны гипофиза. После чего требуется лютеотропное воздействие как ЛГ, так и пролактина, причем пролактин особенно необходим, начиная с 30 дня. Сходство между метэструсом у беременных и небеременных заставляет усомниться в том, что именно воздействие эмбрионов служит сигналом к продолжению секреции прогестерона у сук. Такая ситуация резко отличается от наблюдаемой у многих других домашних животных, у которых присутствие эмбрионов обеспечивает сохранение желтых тел и продолжение секреции прогестерона в течение по крайней мере первой половины беременности. Причем сохранение желтых тел обеспечивается в результате угнетения синтеза простагландинов в матке, и поэтому даже однократная инъекция простагландина вызывает прерывание беременности. У сук такой механизм отсутствует, это демонстрируется тем фактом, что гистерэктомия не оказывает влияния на продолжительность метэструса. Кроме того, желтые тела у собак относительно устойчивы к воздействию простагландинов. Однократная инъекция вызывает у собак временное снижение концентрации прогестерона, а для длительного и существенного уменьшения уровня прогестерона, необходимого для прерывания беременности, требуется повторное введение простагландинов. Как правило, такие инъекции переносятся очень плохо и сопровождаются рвотой и диареей. Установление роли пролактина для поддержания желтых тел после 30 дня привело к многочисленным попыткам искусственно сократить продолжительность метэструса и индуцировать эструс назначением стимуляторов дофамина (бромокриптина, каберголина, метерголина), применение которых рассмотрено в 16 главе. РодыСнижение концентрации прогестерона перед родами наблюдается у большинства домашних животных и объясняется угнетением у беременной самки надпочечниковой секреции стероидных гормонов, что приводит к выработке эстрогена и простагландина эндометрием. У сук родам предшествует резкое падение концентрации прогестерона с 30 нмоль/л (10 нг/мл) до < 6 нмоль/л (< 2 нг/мл) в плазме крови; это, как сейчас принято считать, играет роль пускового механизма родов. Данный факт подтверждается и тем, что назначение прогестерона вызывает задержку родов, а препараты (например, эпостан), блокирующие его секрецию, — стимуляцию. Резкое снижение концентрации прогестерона служит индикатором приближения родов. Концентрация > 6 нмоль/л (> 2 нг/мл) свидетельствует о том, что роды не начнутся в ближайшие 14 часов. Эндокринные тесты на беременностьКак указано выше, пик концентрации прогестерона наблюдается приблизительно на 30 день, после чего она постепенно уменьшается, достигая к концу беременности 30 нмоль/л (10 нг/мл). Градиент снижения концентрации прогестерона после 30 дня у беременных сук практически такой же, как у небеременных. Понятно, что измерение концентрации этого гормона в целях диагностики беременности не имеет смысла. Другими важными для беременности гормонами являются эстрогены. У многих видов, вероятно, в том числе и у собак, синтез эстрогенов происходит в трофобластах эмбрионов. Анализ мочи на содержание сульфата эстрона (основного метаболита эстрадиола), проведенный через 3 недели после вязки, т. е. в период имплантации, теоретически может использоваться для диагностики беременности, однако данный метод не нашел применения. На более поздних сроках в плазме крови определяется эстрадиол, но его концентрация не выше, чем в фазу метэструса, поэтому тесты на эстрадиол не применяются на этой стадии. Содержание белков, таких, как фибриноген, в плазме крови во время так называемой протеиновой фазы повышается на 30–40 день беременности, причем данное явление не характерно для соответствующей стадии метэструса у небеременных сук. Повышение концентрации фибриногена в плазме измеряют с помощью иммунологических методов, а также с помощью существующего готового теста на беременность, хотя с распространением ультразвуковых методов необходимость в таком тесте постепенно снижается. АНЭСТРУС При наличии беременности данную фазу можно определить как интервал между родами и проэструсом, во время которого обычно наблюдается лактация. При отсутствии беременности анэструсом называют интервал между окончанием лютеиновой фазы и началом проэструса. На практике установить окончание лютеиновой фазы достаточно сложно по причине крайне низкого уровня концентрации прогестерона в этот период: обычно он не превышает 3 нмоль/л (1 нг/мл) к 90 дню после пика ЛГ (т. е. анэструс длится по крайней мере 60 дней до начала следующего проэструса). Концентрации эстрадиола и ЛГ изменяются в течение анэструса, однако эти изменения незначительны, и для их выявления требуются многократные анализы крови. Недавними исследованиями установлено, что, несмотря на наличие маленьких фолликулов в период анэструса, они не чувствительны ни к эндогенной, ни к экзогенной гормональной стимуляции. Вероятно, данное обстоятельство связано с рецепторами, но независимо от причины именно это является одним из факторов, вызывающих затруднения в индуцировании фертильного эструса у сук. Относительно высокая концентрация эстрадиола отмечается в течение 10–20 дней до появления внешних проявлений проэструса, что свидетельствует о довольно продолжительном периоде выхода из анэструса. Механизм, запускающий изменения, вероятно, включает активизацию секреции ЛГ, наблюдаемую у некоторых других видов на соответствующей стадии репродуктивного цикла, — например, непосредственно перед переходом от анэструса к эструсу у видов, имеющих сезонный характер размножения, таких, как овцы. Брачный период овец связан с продолжительностью светового дня, обеспечивающего повышение активности гипофиза, и, следовательно, яичников, однако половой цикл домашней собаки не зависит от изменений продолжительности светового дня. Изменения секреции пролактина могут послужить основанием для других объяснений анэструса у сук. Так, концентрация пролактина повышается в период метэструса и в начале анэструса, особенно при беременности. Пролактин обеспечивает лактацию в период анэструса у собак, как и у других видов (свиноматок, кобыл). Этот гормон вызывает снижение чувствительности яичников к гонадотропинам у других видов животных, однако у сук не выявлена последовательная временная корреляция между снижением концентрации пролактина и повышением уровня ЛГ в начале проэструса. При назначении агонистов дофамина (препаратов, подавляющих секрецию пролактина) на стадии метэструса или анэструса концентрация пролактина в плазме крови снижается, соответственно сокращается анэструс. ЗАКЛЮЧЕНИЕ В заключение следует отметить, что влияние гормонов на половой цикл домашней собаки почти такое же, как и у других домашних животных, а основное различие заключается в скорости перехода от одной фазы к другой. Для выявления характерных признаков этих фаз в главе приведены сведения об основных методиках, позволяющих ветеринарному врачу определить время наступления фертильного периода. Кроме того, в данной главе имеется информация, касающаяся процедур, позволяющих воздействовать на репродуктивный цикл суки. Знакомство с физиологией нормального цикла обеспечит понимание методов, применяемых для такого воздействия. ЛИТЕРАТУРА Concannon P. W., Morton D. B. and Weir B. J. (1989) Dog and cat reproduction, contraception and artificial insemination. Journal of Reproduction and Fertility, Supplement 39. Concannon P. W., England G. C. W., Verstegen J. P. and Russell H. A. (1993) Fertility and infertility in dogs cats and other carnivores. Journal of Reproduction and Fertility, Supplement 47. England G. C. W., Allen W. E. and Porter D. J. (1989) A comparison of radioimmunoassay with qualitative and quantitative enzyme-linked immunoassay for plasma progesterone detection in bitches. Veterinary Record 125, 107–108. Jeffcoate I. A. and Lindsay F. E. F.(1989) Ovulation detection and timing of insemination based on hormone concentrations, vaginal cytology and the endoscopic appearance of the vagina in domestic bitches. Journal of Reproduction and Fertility, Suppl. 39, 277–287. Lindsay F. E. F. (1983) The normal endoscopic appearance of the caudal reproductive tract of the cyclic and non-cyclic bitch: post-uterine endoscopy. Journal of Small Animal Practice 24, 1–15. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||