|
||||
|
|
РАЗДЕЛ 1. Отбор животных по поведенческим признакамОбсуждаемые в данном разделе вопросы касаются в основном поведенческих признаков волка, так как именно он был для нас модельным видом. Естественно, что для исследования проблемы реинтродукции в природу выращенных в неволе других видов крупных хищных млекопитающих придется рассматривать другой спектр поведенческих элементов и, соответственно, производить отбор на основе признаков их сформированности. Глава 1.1. Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослыхПоведенческие реакции, сформированные к моменту рождения незрелорождающихся животных и способствующие их первому контакту с окружающей средой, довольно немногочисленны и, судя по литературным данным (Уждавини, 1958; Уждавини, Шепелова, 1966; Слоним, 1967, 1976; Хайнд, 1975; Бадридзе, 1982, 1983 б, 1985, 1987 а, б; Harlow amp; Zimmerman, 1959 и др.), довольно хорошо изучены. Они «как бы пригнаны к условиям существования сразу же после рождения» (Уждавини, Шепелова, 1966, с.106). О подготовленности реакций новорожденных к экологии вида писал и академик П. К. Анохин (1962). Опыты, проведенные в лаборатории А. Д. Слонима (1976), показали, что наиболее выражены, запрограммированные в эмбриональном периоде реакции на кормящую мать. Авторы установили, что можно не только наблюдать врожденные реакции на мать как на внешний фактор, но и моделировать эти внешние факторы. Подобное моделирование проводили и мы (Бадридзе, 1983 а, б; 1 985 1987 а, б). Такой подход способствует выявлению тех раздражителей, на которые возникают конкретные реакции новорожденного животного, что позволяет заполнить пробелы в изучении экологии неонатального периода тех видов, условия обитания которых не дают или ограничивают возможность исследований. Даже в таких значительных работах, как исследования Фокса (Fox, 1971) и Мича (Mech, 1970), касающихся поведения и экологии представителей семейства Canidae и, в частности, волка, нет каких-либо существенных данных об экологической адекватности поведения новорожденных животных. Это можно объяснить большой сложностью наблюдений в природе за контактом новорожденных животных (особенно рождающихся в логове) с матерью и остальной внешней средой, в ответ на воздействие которых развиваются строго определенные врожденные реакции. В лабораторных условиях Фокс (Fox, 1964, 1971) детально исследовал и описал неонатальные реакции Canidae и, в том числе, волка. К сожалению, ни в одной из упомянутых работ нет данных о том, какую роль играет полноценная реализация неонатальных реакций для формирования нормального поведения в постлактационном периоде. Поэтому в данной главе мы попытаемся показать значение полноценной реализации врожденных поведенческих элементов, связанных с процессом питания новорожденных хищных млекопитающих для дальнейшего формирования поведения в пределах нормы. Материал и методикаЭксперименты проводили на 96 новорожденных волчатах, 67 щенках собак (далее — щенки), 30 новорожденных котятах. 10 волчат, 15 щенков и 10 котят были взяты у матери в момент рождения. Возраст отнятия от матери 37 волчат и 15 щенков составлял до 40 часов, а 49 волчат и 37 щенков — 7—10 дней. Всех животных вскармливали с помощью бутылок с соской. Во время кормления 20 волчат и 15 щенков (I контрольная группа) на бутылку с соской надевали диск, диаметр которого составлял 200 мм (рис. 2А), чтобы у новорожденных была возможность упираться лапами во время сосания. Прокол в соске был таков, чтобы не позволять молоку вытекать без сосательных усилий. 10 волчат, 15 щенков и 10 котят (I экспериментальная группа) с момента рождения кормили из сосок без диска, тем самым лишая их возможности упираться во что-либо передними лапками. Остальных котят до окончания лактационного периода вскармливала мать. Во всех случаях у основания бутылки было проделано отверстие диаметром 1 мм для уравнивания давления во время сосания внутри бутылки с атмосферным. Значение этого в свое время было описано (например, Бадридзе, 1987 а, б). 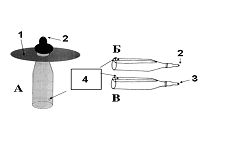 Рисунок 2. Типы применяемых сосок (А, Б, В). 1. Диск; 2. Прокол; 3. Отверстие — 2мм; 4. Отверстия. При вскармливании животных II экспериментальной группы (10 волчат и 10 щенков, отнятых у матери в возрасте 6—40 часов) также применяли два типа сосок: I тип — с проколом, который не позволял молоку вытекать без сосательных усилий, тем самым приближая процесс сосания к естественному (рис. 2Б) и II тип — с отверстием диаметром 2 мм (рис. 2В). При сосании соски с большим отверстием детенышам не приходилось прилагать значительных усилий, и желудок у них наполнялся после двух-четырех сосательных движений. 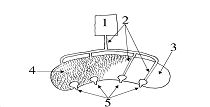 Рисунок 3. Приспособление, имитирующее тело матери. 1-резервуар с молоком; 2-трубки, проводящие молоко; 3-резиновый баллон с водой 36–38 °C; 4-кроличья шкурка; 5-искусственные сосцы. Остальных животных (II контрольная группа, 56 волчат и 27 щенков) вскармливали с помощью сконструированной нами установки, имитирующей тело матери. Установка представляла собой покрытый кроличьими шкурками изогнутый дугой резиновый баллон (рис. 3). Сосцы прикреплялись к баллону со стороны меньшего радиуса. Они соединялись мягкими пластиковыми трубками, пронизывающими баллон. В полость баллона заливалась теплая вода (36–38 градусов), которая нагревала находящееся в трубках молоко. Пастеризованное молоко для вскармливания щенков и волчат приготавливали из молочного порошка, имитируя концентрацию собачьего молока (Барабаш — Никифоров, Формозов, 1963). В результате наблюдения за животными в условиях искусственного вскармливания нами были изучены следующие вопросы: Влияние полноценной реализации «реакции массажа лапками молочной железы» во время сосания на формирование психически уравновешенного взрослого животного. Влияние скорости поступления молока во время сосания в первую неделю жизни на дальнейшее формирование порога сенсорного насыщения. Полученные результаты и их обсуждениеВ свое время мы показали, что «реакция массажа лапками молочной железы» во время сосания в постлактационный период трансформируется в манипуляторную активность при разделке пищи (Бадридзе, 1982, 1983 б, 1985, 1987 а, б. Рис. 4 А, Б). 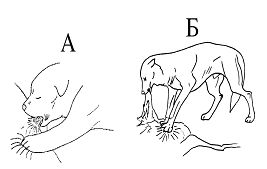 Рисунок 4. Функциональная трансформация реакции массажа лапками молочной железы во время сосания. А — лактационный период; Б — постлактационный период. «Реакция массажа лапками молочной железы» во время сосания представляет собой результат последовательного сокращения разгибательных и сгибательных мышц передних конечностей. В наших экспериментах, когда животных лишали возможности упора лапками во что-нибудь (I экспериментальная группа — 15 щенков, 10 волчат и 10 котят), у них появлялось тоническое напряжение передних конечностей. Это выражалось в растопыривании пальцев и развитии одновременного напряжения сгибательных и разгибательных мышц. Подобное состояние продолжалось до окончания акта сосания. Те новорожденные (I и II контрольные группы), у которых постоянно была возможность опираться на что-либо передними лапками (бутылочка с диском, либо прибор, имитирующий тело матери), осуществлялась нормальная «реакция массажа лапками», т. е. последовательное сокращение реципрокных мышц передних конечностей. В возрасте, когда хищники переходят на полный мясной рацион (40–50 дней), животным начали давать по крупному куску мяса. При первом же его предъявлении все животные контрольных групп начинали его разделывать причем с первого же момента все манипуляционные движения лап были полностью координированы. Иная картина наблюдалась в I экспериментальной группе. Здесь для формирования координированной манипуляторной активности требовалось 3–4 дня. У этих животных заметно проявлялось т. н. «переступание лапами на месте». Параллельно с этим, животные время от времени растопыривали пальцы. Со временем тик ослабевал, но в малозаметной форме сохранялся до конца эксперимента. Кроме того, у всех животных в течение всего периода наблюдений была заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т. н. нормальными) конфликтные ситуации. Соотношение животных контрольной и экспериментальной групп, провоцирующих конфликт, составляло 1/1,25. Волчата и щенки контрольных групп развивались эмоционально нормальными и способны были создать группу с социальными отношениями в пределах нормы. До обсуждения возможных причин подобного различия считаем нужным напомнить данные, приведенные Г. Шепердом (1987), согласно которым импульсация от мышечных и суставных рецепторов поступает через таламус в соответствующую область коры головного мозга. Общеизвестно, что у незрелорождающихся млекопитающих формирование мозговых систем продолжается после рождения и, судя по всему, оптимальный уровень центростремительной импульсации имеет большое значение для их нормального формирования и функционирования. Во время сосания у животных I экспериментальной группы постоянно развивалось тоническое напряжение мышц передних конечностей, что должно было определять центростремительную гиперимпульсацию к соответствующим мозговым структурам. Из сказанного можно предположить, что в период формирования мозговых систем подобная гиперимпульсация может создавать очаги с постоянно высоким уровнем возбуждения, что, в свою очередь, влияет на уровень эмоциональной уравновешенности животного. Из приведенного материала следует, что животные, выращенные при подобных условиях питания в неонатальном периоде, не могут быть пригодными для дальнейшей их реинтродукции. Сравнивая различия в уровне эмоциональной уравновешенности между животными I экспериментальной и контрольными группами, можно заключить, что из-за невозможности нормальной реализации «реакции массажа лапками молочной железы во время сосания» (I экспериментальная группа), развивается одновременное тоническое напряжение всех групп мышц передних конечностей, вызывает возникновение в мозгу очагов постоянно высокого возбуждения. Судя по всему, это и стало причиной эмоциональной неуравновешенности животных этой группы. Животные же контрольных групп имели возможность реализации данной реакции, что, по видимому, и определило нормальное формирование их эмоциональной возбудимости, т. к. у них в мозгу не возникают очаги с постоянно высоким возбуждением. Как было сказано ранее, животных II экспериментальной группы кормили с помощью сосок с отверстием диаметра 2 мм. У волчат и щенков, вскармливаемых из такой соски, желудки наполнялись после нескольких сосательных движений, в течение 0,5 минуты (60–80 мл). Во время подобных экспериментов все животные, несмотря на быстрое наполнение желудка, продолжали проявлять сосательную активность. В свое время нами было показано, что у новорожденных щенят и волчат пищевая мотивация возникает к концу первой недели жизни (Бадридзе, 1987 а, б), а до этого процесс приема пищи осуществляется за счет потребности в реализации сосательной активности. Животные этой группы продолжали принятие чрезмерного количества пищи и после формирования пищевой мотивационной системы. Стремление к перееданию сохраняется в течение всей жизни, что создает постоянные внутригрупповые конфликты на почве конкуренции за пищу. В свое время было показано (Анохин, 1962; Бакурадзе, Чхенкели, 1970 а, б; Бадридзе, 1987 а, б и др.), что включение центральных механизмов головного мозга, тормозящих акт питания, происходит за счет сенсорного насыщения, т. е. центростремительной импульсации, возникающей при растяжении стенок желудка. Анализируя полученный нами материал, можно заключить, что чрезмерный прием пищи в раннем неонатальном периоде может увеличить объем желудка и, следовательно, порог сенсорного (первичного) насыщения, что и определяет развитие относительной гиперфагии. Такие животные активно конкурируют из-за пищи до наступления вторичного насыщения, чем постоянно создают конфликтную ситуацию во время еды. Вместе с тем у них, как правило, не развивается поведение запасания пищи, значение которого будет обсуждаться в следующей главе. Из всего вышесказанного следует, что для нормального развития новорожденных животных, отлученных от матери, решающее значение имеют т. н. «технические» условия при приеме пищи, которые максимально должны быть приближены к естественным. Животные, выращенные в иных условиях, не могут быть пригодны для реинтродукции, т. к. не способны поддерживать в группе полноценные социальные взаимоотношения. Из полученного нами материала можно сделать следующий основной вывод — при искусственном вскармливании хищных млекопитающих для нормального формирования поведения особое значение имеет неонатальный период, который в значительной мере определяет дальнейшее развитие поведения в пределах нормы или патологии. В частности: — для формирования оптимального уровня эмоциональной возбудимости большое значение имеет возможность полноценной реализации реакции «массажа лапками молочной железы во время сосания»; — для исключения высокого порога сенсорного насыщения и, исходя из этого, высокого уровня конкурентных ситуаций в группе во время питания, необходимо условия выкармливания животных максимально приблизить к естественному процессу сосания. Глава 1.2. Поведение запасания пищиЗапасание корма является одним из врожденных видов пищевого поведения, широко распространенного среди животных (Свириденко, 1957; Слоним, 1967; Хайнд, 1975 и др.) Из хищных млекопитающих пищей запасаются собаки, волки, лисицы и др. (Мертц, 1953; Кудактин, 1977; Поярков, 1980; Корытин, 1981; Jeselnik, Brisbin, 1980 и др.) Наиболее полные из имеющихся в литературе сведений по этому вопросу можно найти у Мича (Mech, 1970), однако и их явно недостаточно для исчерпывающего понимания феномена запасания пищи волками. Мич также подчеркивает, что о запасании волками пищи и о том, как это происходит, известно очень мало. Упоминания об этом поведении волка содержатся и в работах Н. А. Зворыкина (1936) и Б. Т. Семенова (1979). В свое время мы изучали закономерность формирования поведения волков при запасании пищи, зависимость способа транспортировки запасаемой пищи от социального статуса особи и специфику памяти на местонахождение кладовых (Бадридзе, 1979, 1987 а, б; Бадридзе и соавт., 1997). К сожалению, мы не встречали в литературных источниках данных, касающихся значения данного вида поведения как для индивида, так и для всей группы в целом и, исходя из этого, для всей популяции. Неизвестно также, может ли существовать естественная патология в формировании данного поведения. Мы имеем в виду естественную патологию, так как причины, из-за которых не формируется поведение запасания пищи при искусственном выкармливании новорожденных животных, уже обсуждались в главе 1.1. Поэтому в данной главе будут освещены вопросы, касающиеся значения поведения запасания для пищевого благополучия индивида, группы и, исходя их этого, для популяции в целом. Изучение данного вопроса должно иметь большое значение для отбора животных по признаку сформированности этого поведения к моменту реинтродукции в природу. Материал и методикаНаблюдения проводили на 78 ручных волках, выращенных в неволе с первых дней жизни до 5-месячного возраста, а на 39 из них — до полового созревания. Помимо этого приведен материал наблюдений за 4 семьями диких волков, приученных к моему присутствию (820 часов наблюдений). Забегая вперед, скажем, что в этой же главе обсудим материал о поведении запасания, полученный на реинтродуцированных нами четырех группах волков. Полученные результаты и их обсуждениеНаблюдения за ручными волками показали, что поведение запасания сформировалось только у 68 волчат. Следует отметить, что из 10 животных, у которых данный вид поведения не сформировался, 8 входили в экспериментальную группу, и причиной несформированности поведения запасания в этом случае была, по-видимому, невозможность осуществления «массажа лапками молочной железы» во время сосания. Два волчонка были представителями контрольной группы, порог сенсорного насыщения в которой сформировался в пределах нормы. Исходя из сказанного, причины несформированности данного поведения у этих животных надо искать во врожденном дефекте, природа которого нам пока непонятна. Мы учитываем этот факт для дальнейшего обсуждения вопроса значимости поведения запасания пищи у волков. В свое время мы установили (Бадридзе, 1987 а, б), что при достаточном количестве добычи, волки после еды 3–4 раза прячут пищу. Надо отметить, что каждый волк запасает от 0,5 до 3, а иногда до 6 кг мяса (последнее зарегистрировано только 4 раза). Средний же вес запаса составлял около 2 кг без учета указанного максимального количества (всего проведено замеров веса содержимого 560 кладовых). Было определено среднее количество особей в группе дикоживущих волков, которое близко к 6 (проведены учеты 83 стай волков; учитывались только взрослые и переярки, т. к. количество запасаемой волчатами пищи менялось в зависимости от их возраста). Если принять число 6 как среднее количество индивидов в стае, то при достаточных размерах жертвы можно представить приблизительное количество запасаемой пищи, которое составит около 36 кг при 3-кратной реализации поведения запасания каждым индивидом. Наблюдения за дикими и реинтродуцированными нами волками показали, что из остатков их трапезы другие плотоядные подбирают, в первую очередь, недоеденные куски, оставшиеся на поверхности. Нанесенные ими потери невелики. В интервалах между охотой волки часто наведываются к запасенной пище. Наши наблюдения в этом плане вполне соответствуют приведенным в литературе данным, которые убедительно показывают, что волки в неблагоприятные для охоты периоды используют кладовые, расположенные в районе добывания пищи (например: Поярков, 1980). Необходимо отметить, что, когда прибылым исполняется 3 месяца, родители в интервале между удачной охотой часто приводят волчат к месту предыдущей трапезы, где они остаются в течение нескольких дней и полностью утилизируют все запасы. Подобное мы постоянно отмечали, наблюдая за дикими и реинтродуцированными нами волками. Даже если учесть, что 3—4-месячные волчата за счет мышкования и ловли насекомых могут на 50 % обеспечить себя пищей (о чем пойдет речь во втором разделе), то можно представить значение запасания пищи взрослыми индивидами как фактора, обеспечивающего волчатам максимально возможную стабильность кормовой базы. Естественно возникает вопрос — что же происходит, когда размер добычи недостаточен для запасания? Наблюдения показали, что, если добычи достаточно для одноразового полного насыщения стаи, но недостаточно для запасания, количество попыток охоты возрастает. Однако это вовсе не гарантирует сокращение интервала между удачными охотами. В тех случаях, когда размер добычи позволяет волкам полностью насытиться, но ее недостаточно для запасания, новая охота начинается спустя 5–7 часов. В случае крупной добычи, когда поведение запасания реализуется в достаточной мере, охота начинается через 2–4 суток. Эти данные приведены в таблице № 2. Необходимо отметить, что у взрослых и переярков количество актов запасания снижается, как только прибылым исполняется 7 месяцев, после чего последние сами интенсивно запасают пищу. А после того, как прибылым исполняется 9 месяцев и они начинают участвовать в охоте взрослых, старшие запасают не более 1 раза. 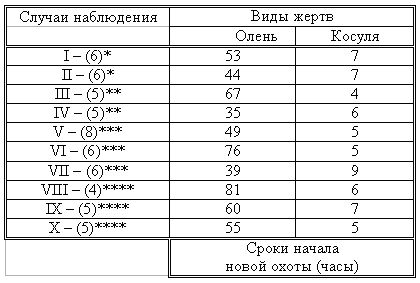 Таблица 2. Зависимость времени начала новой охоты, от размера добытой жертвы. В скобках обозначено количество членов семьи, включая переярков. Таким образом, уровень поведения запасания пищи максимально возрастает в период, когда необходимо обеспечить стабильную кормовую базу для прибылых. В основном это происходит до выхода прибылых из ювенильного возраста. Об интенсивном запасании пищи волками в период выкармливания потомства пишет В. Козлов (1964). Полученный нами материал свидетельствует о том, что поведение запасания пищи представляет собой форму поведения, которая в определенной мере способна обеспечить прибылое потомство стабильной кормовой базой. Поскольку взрослым волкам приходится обеспечивать пищей довольно многочисленное потомство, которое с 3-месячного возраста потребляет почти столько же мяса, сколько взрослые волки, не участвуя при этом в охоте, этот период, на наш взгляд, следует отнести к наиболее трудным в жизни группы. Взвешивая реинтродуцированных нами волков после полного насыщения добытой ими пищи и неоднократной реализации поведения запасания, мы определили, что чистый вес съеденного мяса составлял в среднем 3,5 кг. Если учесть, что каждый волк запасает от 6 до 8 кг, то присутствие в группе одного не запасающего индивида может нанести значительный урон в обеспечении прибылых кормом. Из этого следует, что у животных, отобранных для дальнейшей реинтродукции в природу, поведение запасания пищи должно быть полностью сформировано. Исследовав принципы формирования агрессивности волчат в ювенильном периоде при их 50 %-й пищевой депривации, мы пришли к заключению, что постоянное недоедание меняет приоритеты психологического комфорта (см. Овсянников, Бадридзе, 1989), вследствие чего агрессивное поведение не ритуализируется (Бадридзе, 1997). Вряд ли стоит обсуждать значение ритуализации агрессивного поведения. Итак, феномен запасания пищи у волков представляет собой генетически детерминированный поведенческий акт. Он становится интенсивным во время выкармливания прибылого потомства, тем самым обеспечивая максимально возможную стабильность кормовой базы для волчат в ювенильном периоде. В тех же случаях, когда размер добычи не позволяет делать запасы, интервал между насыщением и охотой значительно сокращается. Это повышает уровень как выживаемости прибылого потомства, так и их формирования в пределах психологической нормы. Резюмируя приведенный материал, можно сделать следующие выводы: — поведение запасания пищи приобретает особое значение в период выкармливания прибылого потомства; — невозможность реализации поведения запасания стимулирует сокращение интервалов между попытками охоты; — наличие в группе даже одной особи, у которой не сформировано поведение запасания пищи, отрицательно сказывается на стабильности кормовой базы волчат; они постоянно недоедают, вследствие чего растет их агрессивность на почве пищевой конкуренции; — особи, у которых не формируется поведение запасания пищи, не должны отбираться для реинтродукции в природу. Глава 1.3. Формирование родительского поведенияВыживаемость вида во многом зависит от уровня развития родительского поведения животного. Уровень сформированности данной категории приобретает особое значение в тех случаях, когда речь идет о реинтродукции в природу выращенных в неволе животных. Практика их разведения в неволе показала, что способные к размножению животные часто не выкармливают детенышей из-за несформированности или неполноценно развитого родительского поведения. Поэтому в процессе отбора животных для реинтродукции в природу особое значение имеет возможность определения индивидов, способных не только к размножению, но и, что очень важно, способных ухаживать за прибылым потомством. Обилие литературных данных, касающихся родительской заботы о потомстве, говорит о том, что это один из наиболее изучаемых видов поведения (Хайнд, 1975; Alexander et al, 1986; Beaver, 1981; Berman et al, 1993; Brown et al, 1995; Capitanio et al, 1984, 1985; Champoux et al, 1992; Clark et al, 1986; Coe, 1990; Fleming, Sarker, 1990; Frankova, 1981; Gonzalez, Deis, 1990; Hector et al, 1989; Hnida, 1985; Insel, 1990; Londei et al, 1995; Numan, 1990; Rosenblatt, 1990; Rosenblatt et al, 1988; Mann et al, 1991). Однако мы не смогли найти ни одной работы, касающейся онтогенеза родительского поведения волка, и, тем более, проблемы прогнозирования вероятности его полноценного формирования. Последнее, как уже отмечалось, крайне важно при отборе животных для реинтродукции в природу. Материал и методикаНаблюдения и эксперименты проводили на 39 волках с 5-месячного возраста до половой зрелости и позже. Чтобы определить, в каком возрасте у волчат проявляются первые элементы родительского поведения, им один раз в месяц предъявляли щенков в возрасте от нескольких часов и до 2 месяцев, и регистрировали реакцию на них. Параллельно с этим волчатам предъявляли также куски материи, пропитанные запахом щенков и — время от времени — магнитную запись их скуления. С годовалого возраста всем волкам предъявляли новорожденных волчат и щенков как одновременно, так и в отдельности. Параллельно велось наблюдение за взаимоотношениями 8 выросших в неволе волчиц и самцов с собственными щенками, а также за взаимоотношениями дико живущих переярков с младшими сибсами. Полученные результаты и их обсуждениеЭксперименты показали, что при предъявлении новорожденных щенков 5-месячным волчатам, у которых не сформировано хищническое поведение, у последних развивается типичное исследовательское поведение с «пробой на зуб». У волчат же, имеющих опыт охоты, запускается типично охотничье поведение. Во всех случаях у волчат с охотничьим поведением щенков отбирали, не давая убить. С 8-месячного возраста у 30 волчат по предъявлении им щенков развивалась четкая исследовательская активность без «пробы на зуб», которая сопровождалась вилянием хвоста и поскуливанием. В течение 1,5–2 минут описанная реакция усиливалась, и волчата начинали охранять щенков от людей, но не от членов своей группы. В дальнейшем, при непосредственном контакте со щенками (в лежачем положении) у волчат, независимо от их пола, развивалась реакция «подставления для сосания», что выражалось прекращением движений тела и поднятием бедра. Вместе с тем, волчата часто вылизывали брюхо и гениталии щенков, что способствовало мочеиспусканию и дефекации. Во всех случаях скуление щенков вызывало сильное возбуждение волчат, которые начинали скулить, интенсивно обнюхивать щенков и их вылизывать. Через 4–5 часов после предъявления щенков волчата теряли к ним интерес и оставляли их. Время от времени интерес к щенкам возобновлялся, волчата подходили к ним и обнюхивали их в течение 1,5–2 минут. В это время они уже не охраняли щенков от людей. К 10—11-месячному возрасту реакция ухаживания волчат за щенками становилась прочной и постоянной. В этом возрасте они начинали давать месячным щенкам отрыжку, независимо от того, выпрашивали они ее или нет. У 9 волчат в течение всего описанного периода исследований при предъявлении щенков развивалась исключительно исследовательская активность с «пробой на зуб». Эксперименты показали: что при одновременном предъявлении новорожденных щенят и волчат остальным 30 годовалым волкам, они, независимо от пола, начинали интенсивно обнюхивать их, а спустя 10–20 секунд брали волчат в пасть и относили в «укромное место», после чего родительское поведение развивалось в полной мере. Только после этого у волков развивалась агрессивная реакция на щенков. У остальных 9 волков развивалась такая же исследовательская активность, как и в 8-месячном возрасте, с той лишь разницей, что они исследовали волчат почти в 2 раза дольше, чем щенков (волчат около 60 секунд, щенков — около 30). Через неделю после «усыновления» волчат годовалыми волками, у одной из 22 самок, волчицы «Рухи» началась лактация, что полностью исключило их искусственное выкармливание. Для иллюстрации того, насколько сильна у волков мотивация ухаживания за новорожденными, приведу такой случай. Волчица «Ники» родила пятерых волчат. В первые три дня никогда не рожавшая волчица «Хешке», которая имела в группе низкий статус, постоянно пыталась залезть в логово к волчатам, но получала яростный отпор «Ники». В течение этих дней у «Хешке» заметно вздулись сосцы, а на четвертый день она, яростно агрессивная, ворвалась в логово и выгнала «Ники». Последующие три дня «Хешке» не покидала логово, не подпуская к нему «Ники». Очевидно, у нее было достаточно молока для кормления волчат. На четвертый день она вышла из логова и улеглась перед ним. Воспользовавшись этим, «Ники» вбежала в логово и легла с волчатами, которые принялись сосать ее. Приблизительно через три часа «Хешке» с рычанием снова выгнала ее из логова и начала кормить волчат. С этого момента волчицы сменяли друг друга примерно три раза в сутки. Так продолжалось до окончания лактационного периода. Следует отметить, что у всех самцов положительная реакция на щенков формировалась на 1–1,5 недели позже, чем у самок, хотя интенсивность поведенческой реакции родительской заботы развивалась у них одинаково. Как было сказано выше, волчатам с 5-месячного возраста предъявляли куски материи, пропитанные запахом щенков, а также воспроизводили магнитную запись скуления щенков. Эксперименты показали, что у 5-месячных волчат в ответ на предъявление запаха новорожденных щенков развивалась исключительно исследовательская активность, выражавшаяся в интенсивном обнюхивании источника запаха. Спустя 15–20 секунд интерес к этому пропадал. При предъявлении же магнитной записи у волчат запускалась ориентировочная реакция. Затем, после определения местонахождения звука у них развивалась хорошо выраженная исследовательская активность, которая проявлялась в интенсивном обнюхивании и касании лапой, у некоторых волчат развивалась и реакция «пробы на зуб». Исследования проводились в несколько серий продолжительностью 40–60 секунд с интервалами в 15–40 секунд. У тех же 30 волчат, у которых в возрасте 8 месяцев развивалась положительная реакция на предъявляемых щенков, в ответ на предъявление запаха и звука новорожденных щенков развивалась исследовательская активность, выражавшаяся в скулении и интенсивном вилянии хвостом. У этих волчат ни в одном случае не развивалась реакция «пробы на зуб». При одновременном предъявлении и запахов, и звуков новорожденных щенков волкам в возрасте 1 года и старше, у них также развивалась исследовательская активность со скулением и вилянием хвостом. Вместе с тем, наблюдения показали, что запах волчат исследуется волками почти в 2 раза дольше, чем запах щенков, в то время как источник звука при воспроизведении скуления волчат и щенков исследуется ими одинаковое время (таблица 3). В течение 3 месяцев мы в естественных условиях наблюдали, как два переярка выкармливали месячных волчат, мать которых была убита егерями (остался ли в живых самец, неизвестно). Переярки, которые, судя по всему, были членами той же семьи, выполняли функцию волчицы в течение всего периода наблюдений. 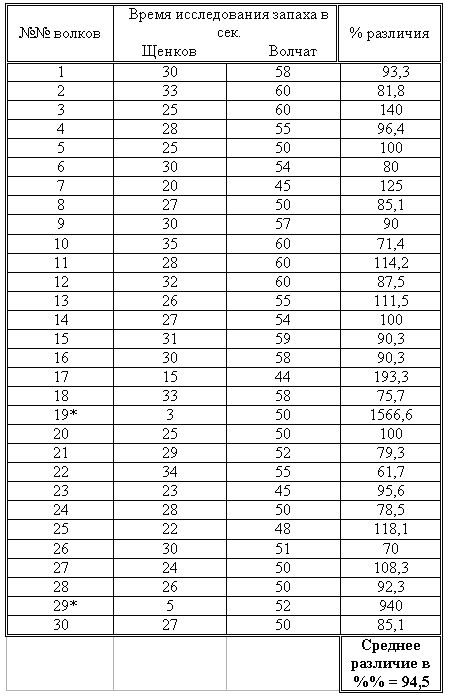 Таблица 3. Продолжительность исследования годовалыми волками запаха волчат и щенков. Исходя из полученных нами результатов, процесс формирования родительского поведения у волков начинается с 8 месяцев и заканчивается к годовалому возрасту, т. е. к моменту появления в семье новой генерации. Судя по всему, данный вид поведения запускается в результате восприятия детенышей. Интересно, что положительная реакция в одинаковой мере возникает у волков при предъявлении им как волчат, так и щенков, однако при наличии выбора предпочтение отдается своему виду. По-видимому, что именно видовой запах имеет решающее значение для подобного предпочтения. Визуально определяемые признаки и звуковые сигналы новорожденных волчат и щенков, по всей видимости, не имеют для волков различимой видовой специфики, они несут только один признак — признак инфантильности. Вместе с тем, комплексное восприятие инфантильных признаков может вызвать у животных с полноценно развитым родительским инстинктом физиологические сдвиги настолько сильные, что они могут провоцировать лактацию, в ряде случаев, даже у неполовозрелых самок. Необходимо отметить, что из тех 9 волков, у которых не сформировалось родительское поведение, четверо были взяты из Московского, а трое — из Тбилисского зоопарков, остальные же два волчонка были отняты охотниками у диких волков. Из этих животных только две волчицы родили детенышей, но в тот же день бросили их, из-за чего волчат пришлось выкармливать искусственным образом. Причины данного явления нам определить трудно, но, независимо от этого, подобные животные не могут быть отобраны для реинтродукции в природу, так как не способны обеспечить высокую выживаемость прибылого потомства и, соответственно, выживаемость реинтродуцированной популяции. В то же время, для определения пригодности животных к реинтродукции в природу, нет необходимости ждать их размножения, так как сформированность данного признака можно определить задолго до половой зрелости животного. Резюмируя приведенный материал, можно сделать следующие выводы: — у волков родительский инстинкт начинает проявляться с 8 месяцев и полностью формируется к годовалому возрасту, то есть к моменту появления в семье нового потомства; — у индивидов со сформированным родительским инстинктом данный вид поведения запускается за счет восприятия визуальных, обонятельных, звуковых и инфантильных признаков; — животные с неразвитым родительским инстинктом не могут быть отобраны для реинтродукции в природу; — в процессе отбора животных для реинтродукции в природу их можно задолго до половой зрелости тестировать на развитость родительского инстинкта с помощью комплексного или раздельного предъявления раздражителей, несущих инфантильные признаки. Глава 1.4. Отбор животных по признаку уровня рассудочной деятельностиЗначение изучения рассудочной деятельности животных трудно переоценить как с научной, так и с этической точки зрения. Большой вклад в изучение этой проблемы внес Л. В. Крушинский и его сотрудники (Крушинский, 1958, 1960, 1968, 1977; Крушинский и соавт., 1965, 1980, 1981, 1985). В этих работах представлены результаты изучения указанного феномена на нескольких таксономических группах животных, разного филогенетического уровня и убедительно показана высокая способность многих видов животных к элементарной рассудочной деятельности. Объектом опытов Л. В. Крушинского был также и волк, большая адаптивная способность которого дала ему возможность не только выжить в условиях многовекового преследования человеком, но и в большей мере, чем другим животным, сохранить свой ареал. Помимо этого, высокий уровень элементарной рассудочной деятельности волка определяет и совершенство методов его охоты на крупную жертву. Одним из проявлений феномена рассудочной деятельности является способность животного к экстраполяции направления движения раздражителя, исчезающего из поля зрения, которая, по формулировке Л. В. Крушинского (1977), дает ему возможность выносить известную функцию на отрезке за его пределы. Изучив эту способность, автор показал, что успех решения экстраполяционных задач при их многократном предъявлении в ряде случаев снижается в результате трудностей, вызванных резким возбуждением мозга. Естественно, возникает вопрос — каким же образом решают волки экстраполяционные задачи, многократно возникающие во время охоты, когда уровень эмоционального возбуждения у них и так довольно высок? Весьма значительной представляется проблема взаимосвязи рассудочной деятельности и обучения, важность которой в свое время отметил Л. В. Крушинский (1977). Изучение данной проблемы имеет, помимо фундаментального, и прикладное значение, так как позволит определить те способы выращивания животных в неволе, которые обеспечат формирование характерного для данного вида максимального уровня рассудочной деятельности. Решение данной проблемы (наряду с другими) позволит полноценно подготовить выращенных в неволе крупных хищных млекопитающих к реинтродукции в природу. К сожалению, в литературе эта проблема до сих пор не обсуждалась. В данной главе мы попытаемся осветить вопросы, касающиеся проблемы формирования у выращиваемых в неволе животных характерного для данного вида максимального уровня рассудочной деятельности. Материал и методикаЭксперименты и наблюдения проводили как в вольерах (100 м2), так и в полевых условиях на 78 волках до 5-месячного возраста, из них на 39 — до полового созревания. Первую группу — экспериментальную (39 особей) содержали в вольере, где одновременно находились до 6 особей. В вольере были установлены крупные непрозрачные предметы: валуны разных размеров, деревянные ширмы (120x90 см), насыпи и завалы из бревен. Все предметы были установлены так, чтобы звери могли свободно перемещаться среди них. Часть рациона волчат состояла из живых крыс и кроликов (рис 5А). Вторую группу — контрольную (39 особей) содержали в вольере, где также одновременно находились до 6 особей. Здесь отсутствовали крупные предметы, за исключением двух ширм (120 на 90), вплотную придвинутых к стене вольера с той стороны, откуда в поле зрения животных никто никогда не появлялся. За эти ширмы не могли проникнуть ни волки, ни крысы, ни кролики, которых скармливали этим хищникам так же, как животным экспериментальной группы (рис 5Б). 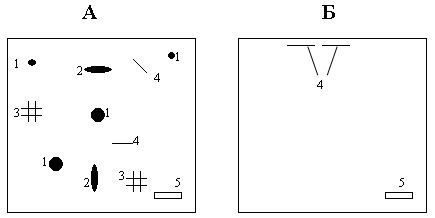 Рисунок 5. Условия содержания экспериментальной (А) и контрольной (Б) групп. 1 — валуны; 2 — насыпи; 3 — завалы из бревен; 4 — ширмы; 5 — поилки. В 7-месячном возрасте животных обеих групп (экспериментальная n=25, контрольная n=14) по одному разу тестировали на экстраполяционной установке Крушинского (1960). Во всех случаях за 7 дней до тестирования эта установка собиралась из ширм, находящихся в вольере, где и происходили опыты. Основные эксперименты на экстраполяционной установке проводили на голодных волках годовалого возраста. В процессе тестирования особое внимание уделяли динамике успешности решения задач при их многократном предъявлении. В течение опытного дня каждому волку предъявляли 30 экстраполяционных задач с интервалом в одну минуту. Во всех случаях вес приманки составлял 50 граммов. При тестировании в вольере находился один зверь. Общее число предъявлений теста каждому хищнику составило 300. По окончании тестирования животных контрольной группы в их вольере создавались такие же условия, как и в экспериментальной, то есть среда их обитания обогащалась за счет установки крупных непрозрачных предметов, после чего наблюдали за взаимодействиями животных между собой и с живым кормом. Принцип содержания волчат экспериментальной группы был подсказан наблюдениями за дикими волчатами. Кроме описанных, наблюдения за животными обеих групп проводили и в полевых условиях. Особое внимание обращалось на способность адаптации к среде обитания и на успешность охоты на диких животных. 22 волка экспериментальной группы вывозили в поле с возраста 4 месяцев, а 14 животных из контрольной и 3 — из экспериментальной групп — в возрасте не менее 2 лет. Полученные результаты и их обсуждениеНаблюдения показали, что волчата, выращиваемые в неволе, так же, как и дикие, много времени уделяют играм (не менее 1,5 часа в сутки, в среднем около 3-х часов), которые заключаются в возне, борьбе и преследовании друг друга. Преследования включали и конкурентные взаимодействия из-за пищи. Во время этих игр преследуемые волчата использовали всевозможные укрытия: стволы деревьев, завалы, валуны и т. п. Наблюдения за дикими волчатами показали, что в процессе преследования нападающие в раннем возрасте повторяли путь преследуемого волчонка, а позже (в возрасте около 6 месяцев) начинали срезать путь преследования. Во время игры в «догонялки» преследуемый и преследующий часто менялись ролями. Животные из экспериментальной группы (так же, как и дикие) до 6-месячного возраста во время преследования в основном повторяли путь убегающего. Та же картина наблюдалась и в случае предъявления им живых крыс и кроликов. Мы говорим «в основном», так как иногда преследующие срезали путь преследуемого волчонка или жертвы, зашедших за непрозрачные предметы. С возрастом количество таких маневров увеличивалось и к 7-месячному возрасту достигало 80 % (рис. 6). В этом возрасте животных из обеих групп тестировали на экстраполяционной установке. Тестирование показало, что из 25 животных экспериментальной группы задачу при ее первом предъявлении решили 20 (около 80 %). В контрольной группе задачу при первом предъявлении решили 6 из 14 животных (около 42,8 %). 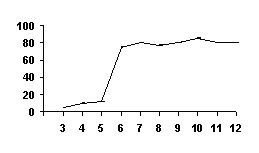 Рисунок 6. Динамика процента случаев срезания пути преследования (обход непрозрачных предметов) сверстников и жертвы, волками экспериментальной группы. X — возраст в месяцах; Y — процент случаев срезания пути преследования; Выборка: n=250 в месяц для каждого преследующего.  Рисунок 7. Процент правильных решений экстраполяционных задач при их одноразовом предъявлении семимесячным (А) и годовалым (Б) волкам экспериментальной (Э) и контрольной (К) групп. 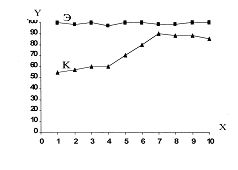 Рисунок 8. Динамика (в процентах — Y) экстраполяционных задач при их многократном предъявлении. Х — экспериментальные дни (30 задач в день с интервалом 1 минута; Э — экспериментальная и К — контрольная группы. Опыты с экспериментальной группой показали, что, независимо от числа предъявлений теста, все животные решали его успешно. Большинство животных контрольной группы при первом предъявлении решало успешно (12 животных из 14; 85,7 %), а при многократном предъявлении успешность их решения снизилась и начала снова повышаться после четвертого дня эксперимента, т. е. после 120-го предъявления задачи. У 11 животных из 14 (около 78,5 %) к 180-ому предъявлению успешность решения задач достигла 90 % (рисунки 7 и 8). Четверо животных контрольной группы в среднем после 11-ого предъявления вообще отказались от решения задач. При попытке предъявления им экстраполяционных задач они начинали беспокоиться и рычать. При начале движения приманки они стали издавать специфический фыркающий лай тревоги. У всех четырех животных сама экспериментальная ситуация вызывала сильное эмоциональное напряжение. Динамика успешного решения многократно предъявляемых экстраполяционных задач показана на рисунке 7. При анализе материала не учтены данные по четырем животным, которые отказались от многократного решения задач. Как отмечалось выше, после окончания тестирования животных контрольной группы, в вольере, где они содержались, устанавливали крупные непрозрачные предметы и в течение 6 месяцев наблюдали динамику количества случаев срезания пути преследования сверстников и живого корма. Наблюдения показали, что в течение всего этого времени процент случаев срезания пути преследования медленно нарастал, но так и не превысил 35 % (рисунок 9), что существенно ниже: чем у животных: выросших в вольере с крупными предметами. 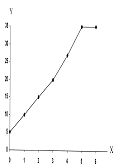 Рисунок 9. Динамика (в месяцах — Х) процента (Y) случаев срезания пути преследования у годовалых волков контрольной группы, после обогащения среды непрозрачными предметами. 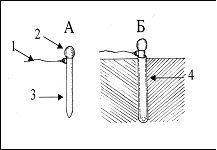 Рисунок 10. Приспособление для ограничения свободы передвижения волка. А — свободный стержень; Б — стержень, вставленный в вбитый в землю цилиндр. 1 — трос; 2 — деревянный набалдашник; 3 — стержень; 4 — вбитый в землю цилиндр. Для того чтобы выяснить, способны ли волки прогнозировать результат своего воздей-ствия на внешний раздражитель, были поставлены эксперименты на 37 животных (24 волка из экспериментальной группы и 13 — из контрольной. Начинали эксперимент на волках в возрасте 7,5 месяцев). С двумя животными (1 из экспериментальной группы, 1 — из контрольной) эксперимент начинали, когда они достигли возраста 1,5 года. Ни один из зверей не был приучен к содержанию на привязи. С 7-месячного возраста каждому подопытному волку к ошейнику пристегивали трос длиной в 3 метра, на противоположном конце которого был прикреплен металлический стержень с деревянным набалдашником (рисунок 10А). После прикрепления троса к ошейнику волкам предоставлялась неограниченная возможность передвижения в течение 5 минут, после чего трос снимали. Через 15 дней, т. е. после полного привыкания к тросу, начинали ставить следующие эксперименты: к ошейнику волка прикрепляли трос, а металлический стержень вставляли в металлический цилиндр, предварительно вбитый в землю (рис. 10Б), чем и ограничивали возможность его передвижения. После этого перед волком на недосягаемом для него расстоянии (4 метра) выкладывали полную порцию пищи и регистрировали его поведение. Подобные эксперименты проводили с интервалом в 3 дня, каждый раз тестируя животное по одному разу в течение 60 минут. По истечении этого времени, т. е. когда волк не мог найти способа освободиться от привязи, пищу убирали, а животное освобождали. После полного привыкания к такой ситуации, т. е. когда волки при предъявлении им пищи внешне переставали на нее реагировать, отворачивались и ложились (после 15–20 тестирований), мы на виду у них доставали стержень из цилиндра, тем самым предоставляя свободу действий. Полуторагодовалых волков тестировали по одному разу каждый день в течение одного часа, не предоставляя свободы. Как уже отмечалось, на двух животных подобные эксперименты начали проводить с полуторагодовалого возраста. Принцип приучения животных к экспериментальной ситуации и сами эксперименты были такими же, как и с остальными животными. Эксперименты с основной группой волков показали, что в течение первых 15 дней все волки полностью привыкали к ношению троса на ошейнике и не реагировали на него. При первых 15–20 тестированиях животные бурно реагировали как на ограничение свободы, так и на невозможность добраться до пищи. Все волки пытались освободиться, пятясь назад, стараясь сбросить ошейник, и время от времени грызли трос. Через 15–20 тестирований все волки после предъявления им пищи в течение 1–1,5 минут отворачивались от нее, ложились и внешне успокаивались. Во всех случаях, когда после этого на виду у волка освобождали трос, вынимая стержень из цилиндра, все они сразу же вскакивали и подбегали к пище. Эксперименты с полуторагодовалыми волками основной группы показали, что при тестировании в этом возрасте животные во время предъявления им теста вновь начинают волноваться. Это выражалось в повышении общей активности, скулении, которое время от времени сменялось рычанием, а также в попытках перекусить трос. Повышенная активность не спадала в течение всего времени тестирования. Во время четвертого тестирования, спустя 15 минут после его начала волк N6 («Малыш»), вновь охваченный волнением, с рычанием подбежал к стержню, державшему трос, схватил его за набалдашник и извлек из цилиндра, после чего моментально выпустил его и бросился к пище. Во время седьмого тестирования тот же волк смог освободиться в течение первых 20 секунд, причем без заметного волнения, и бросился к пище. Все остальные волки эту задачу смогли решить подобным образом при 7—12-ом тестировании. У двух волков, которые подверглись эксперименту в полуторагодовалом возрасте, способность к решению поставленной задачи развивалась по-разному, несмотря на то, что скорость адаптации к условиям эксперимента была практически одинаковой. Волк экспериментальной группы, который в этом возрасте успешно решал экстраполяционные задачи при их многократном предъявлении, при данном тестировании начал решать задачи после 22-го предъявления. Вместе с тем, уровень нервного напряжения у него был такой же, как и у членов основной группы. Волк контрольной группы, у которого уровень нервного напряжения был несравненно выше, предъявленные задачи начал решать только после 85-го тестирования. Необходимо отметить, что полевые исследования, проведенные на обеих группах волков, показали, что у всех животных контрольной группы и у упомянутых трех волков экспериментальной группы процесс адаптации к полевым условиям протекал почти в 2 раза дольше, чем у животных экспериментальной группы. Самым важным нам представляется то, что у всех животных контрольной группы процент успешной охоты никогда не превышал 10 %, т. к. они постоянно повторяли путь преследуемой ими потенциальной жертвы. Это продолжалось в течение всего периода наблюдений (5 месяцев), тогда как у всех волков экспериментальной группы, включая трех упомянутых, с первых же дней успешность охоты на диких животных составила 50 %. Следует отметить, что в течение последующих 6 месяцев процент удачных охот снизился по независящим от волка причинам, которые будут обсуждаться в последней главе, т. к. не могут быть отнесены к проблеме рассудочной деятельности. Надо отметить также, что у животных обеих групп были одинаковые условия для охоты, т. к. в данном районе (детальные сведения будут приведены также в последней главе) за многие годы популяция волка была полностью истреблена, и дикие копытные убегали от наших волков с расстояния всего около 15 метров. Как видно из полученных нами результатов, у диких волчат и волчат экспериментальной группы принципиально одинаково развивается способность обхода непрозрачных предметов в процессе преследования партнера или жертвы. Мы считаем, что нарастание количества подобных обходов непрозрачных предметов в значительной степени определяется положительным подкреплением, вызванным поимкой партнера по игре или жертвы. Все это, по нашему мнению, говорит о том, что у волчат 7-месячного возраста (экспериментальная группа) способность обходить непрозрачные предметы, срезая путь преследования партнера или жертвы, т. е. способность экстраполяции направления движения развивается за счет приобретенного опыта — обучения. Волчата того же возраста из контрольной группы были не способны решать подобные экстраполяционные задачи. Как видно из рисунков 7 и 8, годовалые животные из экспериментальной группы практически с одинаковым успехом решают экстраполяционные задачи как при первом, так и при многократном предъявлении. Волки же контрольной группы, успешно справившись с задачами при первом предъявлении, несравненно хуже решают их при многократном предъявлении. Причины снижения успешности решения экстраполяционных задач в подобных условиях предъявления Л. В. Крушинский объясняет «трудностями, которые возникают в процессе решения задачи, при обучении этого не происходит» (1977, с.53). Далее подчеркивается, что эти «трудности» вызывают резкое возбуждение мозга, что и является причиной снижения критерия достоверности решения экстраполяционных задач. Как отмечалось выше, у 7-месячных волков экспериментальной группы способность срезать путь преследования партнера или жертвы развивается за счет опыта — обучения. Вместе с тем, в указанном возрасте волчата способны успешно решать экстраполяционные задачи, предъявленные с помощью установки Крушинского. Тут закономерно возникают следующие вопросы: — каким образом опыт обхода непрозрачных предметов, полученный при взаимодействии со сверстниками или жертвой, волки применяют в решении задач, предъявленных с помощью экстраполяционной установки? — каким образом 7-месячные волки могут решать предъявляемые им экстраполяционные задачи, если способность к элементарной рассудочной деятельности у этого вида проявляется в годовалом возрасте (учитывая результаты, полученные на контрольной группе)? Для того чтобы ответить на поставленные вопросы, обратимся к рисунку 6, где показана динамика срезания пути преследования сверстника или жертвы. Из рисунка видно, что у волков в возрасте от 3 до 5 месяцев происходит очень медленное нарастание количества адекватных обходов непрозрачных предметов при преследовании сверстника или жертвы. А с 5-месячного возраста количество подобных обходов нарастает взрывообразно (около 20 %) и к 6-месячному возрасту достигает 75 %. Исходя из полученных данных, мы склонны считать, что у волчат в возрасте от 3 до 5 месяцев происходит сравнительно равномерное, медленное накопление опыта адекватного обхода непрозрачных предметов во время преследования сверстников или жертвы. Опыт закрепляется, когда удачный обход заканчивается поимкой партнера или жертвы, то есть получает положительное подкрепление в виде экономии энергетических затрат, быстрого появления преследуемого объекта на более близкой дистанции и т. д. Мы считаем, что количество адекватных обходов у волчат 5-месячного возраста резко возрастает не только благодаря обучению, но и развитию качественных изменений в высшей нервной деятельности. В пользу того, что в данном возрасте мы имеем дело с качественным изменением в высшей нервной деятельности, а не с увеличением способности к обучению, говорит и то, что у четырех- и семимесячных волчат скорость выработки условного пищевого рефлекса (обучения) на различные зрительные раздражители еще существенно не отличается (Р›0,05). Возвращаясь к поставленным выше вопросам, можно заключить, что у волков в возрасте от 5 до 7 месяцев формируется способность переноса ранее приобретенного опыта в новую, но логически схожую ситуацию. Эта способность, видимо, и является качественно новой формой высшей нервной деятельности, развивающейся на данном этапе постнатального онтогенеза. Именно она и обеспечивает решение экстраполяционных задач 7-месячными волками из экспериментальной группы. Исходя из этого, можно было бы допустить, что описанная способность является первым этапом в развитии способности волка к рассудочной деятельности. Второй же этап в развитии данной способности формируется к годовалому возрасту (пример животных контрольной группы). Именно в этом возрасте у волков появляется способность к решению рассудочных задач без всякого предварительного опыта, т. е. как формулирует Л. В. Крушинский (1977): допуская, что в определенных случаях животное при решении рассудочной задачи может применять предшествующий опыт, решение новой задачи происходит на «эвристическом уровне» работы мозга, т. е. за счет озарения — insight («озарение» — подразумевается формулировка Торпа — Thorp, 1958). Мы не можем согласиться с тем, что «мыслящие» животные способны решать новые задачи, если информация хотя бы о нескольких их элементах не дает возможности логического сопоставления с существующим опытом. Именно с этой позиции постараемся разобраться, почему существует разница в успешности многократного решения экстраполяционных задач волками экспериментальной и контрольной групп. В свое время П. К. Анохин писал, что «важная сторона поведения животных и человека — механизмы реализации накопления опыта в свободном поведении, пластическое комбинирование «фрагментов памяти» и построение на этой основе новых форм поведения» (1962, с.10). Говоря об использовании ранее приобретенного опыта в решении новой рассудочной задачи, мы подразумеваем пластическое комбинирование фрагментов образной памяти, в изучение которой огромный вклад внес патриарх грузинской физиологической школы И. С. Бериташвили (1968, Беритов, 1969). Но на каком уровне происходит оперирование этой памятью? Анализируя полученные нами результаты, мы пришли к заключению, что волки контрольной группы могут приобретать «пассивные знания» о неисчезаемости предметов, заходящих за непрозрачные предметы. Подобные «знания» могут возникать за счет наблюдения за движением сверстника, когда тот заходит за другую особь. Аналогичные «знания» могут формироваться в отношении неконтролируемого количества ситуаций и раздражителей. Мы имеем в виду «пассивные знания», т. к. у волков контрольной группы не может формироваться двигательный навык взаимодействия с подобными движущимися объектами. У волков же экспериментальной группы подобный навык формируется и закрепляется в течение длительного времени постнатального онтогенеза. Для дальнейшего обсуждения нашего материала целесообразным представляется обратиться к существующим в литературе данным. И. С. Бериташвили показал, что индивидуально приобретенное поведение в основном регулируется путем репродукции образов жизненно важных объектов, названной им образной психонервной деятельностью (в отличие от специфической психонервной деятельности человека — сознательной). Касаясь феномена сознания, П. В. Симонов (1985) пишет, что в процессе длительной эволюции подсознание возникло как средство защиты сознания от стрессирующих перегрузок, иллюстрируя это подсознательной реализацией двигательных навыков у пианистов, спортсменов и т. д. При обсуждении нашего материала, кроме понятия образной психонервной деятельности как аналогии сознательной деятельности человека, будем использовать понятие процессы, не выходящие на уровень образной психонервной деятельности, как аналогию подсознательного. Возвращаясь к полученным нами данным, считаем возможным утверждать, что волки экспериментальной группы, у которых навык адекватного обхода непрозрачных предметов при преследовании сверстников или жертвы формировался и закреплялся в течение длительного времени, способны многократно решать экстраполяционные задачи за счет прочного предварительного опыта; применение такового не должно вызывать процессы, выходящие на уровень образной психонервной деятельности. Мы склонны считать, что именно подобный механизм решения экстраполяционных задач должен обеспечить сохранение нормального уровня нервного напряжения. Волки же контрольной группы, не имеющие активного и прочного навыка взаимодействия с «исчезающими» движущимися предметами, способны решать экстраполяционные задачи при условии активного оперирования ранее приобретенной информацией. Это активное оперирование, по-видимому, вызывает процессы, выходящие на уровень образной психонервной деятельности, чему должен сопутствовать высокий уровень нервного напряжения, вызывающий срывы при многократном предъявлении экстраполяционных задач. Независимо от того, на каком уровне нервной деятельности животные обеих групп решают экстраполяционные задачи, эти решения достигаются за счет адаптивной реорганизации имеющихся у каждого индивида «знаний», что обеспечивает высокую вероятность нахождения «логических» связей между новой задачей и задачами, решенными в прошлом. К. Г. Прибрам (1982) в своей обзорной работе показал наличие специализаций корковых областей большого мозга как в выборе альтернатив, так и в установлении знакомого контекста, в рамках которого информация может быть обработана. Как мы уже отмечали, годовалые животные из контрольной группы после обогащения среды их обитания в неволе не способны научиться срезать путь преследования партнера или жертвы. Процент этого действия медленно нарастает, но так и не превышает 35 %. Судя по всему, пластическое комбинирование «фрагментов памяти», которое происходит за счет нахождения логических связей между имеющимся опытом и конкретной ситуацией, и построение на этой основе новых форм поведения, способность к которому у волков возникает и развивается в возрасте от 5 до 6 месяцев, требует постоянной тренировки. В ином случае способность к подобной мозговой деятельности может с возрастом затрудняться, вследствие чего высокому нервному напряжению при решении таких задач животные предпочитают большие энергетические затраты. На наш взгляд, это хорошо подтверждает тот факт, что некоторые голодные животные предпочитают отказаться от решения экстраполяционной задачи, дающей пищевое подкрепление. Таким образом они делают выбор между пищевой потребностью и потребностью сохранить нормальный уровень нервного напряжения, тем самым сохраняя «психологический комфорт» (см. Овсянников, Бадридзе, 1989). Обсуждая полученные нами данные о способности волков прогнозировать результат своего воздействия на внешний раздражитель, стоит напомнить некоторые результаты: когда в поле зрения лежащих волков мы освобождали трос, вынимая стержень из вбитого в землю цилиндра, волки сразу вскакивали и подбегали к пище. На наш взгляд, это говорит о том, что у них в течение первых 15–20 дней тестирования вырабатывается опыт возможности свободного передвижения при вынутом из цилиндра стержне. Вместе с тем, до полуторагодовалого возраста у волков должен накапливаться зрительный опыт самого процесса освобождения, т. е. процесса изъятия из цилиндра, вбитого в землю, стержня, который удерживает трос. Доставая стержень из цилиндра, они, вероятно, используют именно этот опыт. Мы склонны считать, что подобные действия волки совершают за счет развитой на данном этапе онтогенеза способности к прогнозированию результата своего воздействия на внешний раздражитель. Сама способность в данном случае реализуется за счет знания, что свободный стержень означает свободу передвижения, а также за счет визуального опыта, связанного с процессом, дающим эту свободу. Мы считаем, что только способность животных оперировать предшествующим опытом дает им возможность прогнозировать результат своего воздействия на внешний раздражитель, а сама способность прогнозировать результат своего воздействия на внешний раздражитель, видимо, является наиболее сложной формой высшей нервной деятельности, развившейся на данном этапе онтогенеза. Эту мысль подтверждают данные, полученные нами (Бадридзе, 1987 а, б) при изучении развития навыков групповой охоты волков: животные в возрасте 1,5–2 лет после, пусть даже случайно приобретенного опыта (который не зависит от действий самих волков) способны загонять жертву в места, где с ней легко справиться, формируя тем самым традицию приема охоты. Тот же феномен мы наблюдали в случае с реинтродуцированными нами в природу волками, о чем пойдет речь ниже. Анализируя описанные результаты, считаем необходимым еще раз напомнить высказывание Л. В. Крушинского и соавторов, что «способность к элементарному мышлению не только дает возможность животному адекватно реагировать на статическое соотношение раздражителей, но и выполнять определенные элементарные «прогнозирующие» реакции в процессе изменения данного соотношения» (1965, с.58). Учитывая полученные нами данные, считаем возможным заключить, что «думающие» животные и, в частности, волк (как вид) при наличии определенного опыта может не только выполнять элементарные «прогнозирующие» реакции при изменении соотношения раздражителей, но и прогнозировать результат целенаправленного изменения им этого соотношения. Это заключение может в определенной степени быть ответом на поставленный Фриджлинком (Frijlink,1977) вопрос: умышленно ли пользуются волки конкретным приемом загона жертвы (оленя). Целесообразно напомнить, что у полуторагодовалых волков, которые до этого возраста были полностью адаптированы к экспериментальной ситуации, в процессе тестирования вновь возникало эмоциональное напряжение. Судя по всему, эмоциональное напряжение возникает именно в тот период, когда у животного начинает формироваться способность к прогнозированию результата целенаправленного изменения соотношения внешних раздражителей. Непосредственной же причиной возникшего нервного напряжения в данной ситуации, как нам представляется, является конфликт между способностью улавливать логическую связь явлений и неумением на данный момент принять решение для изменения соотношения раздражителей. Мы показали, что двухгодовалые волки контрольной группы не способны полноценно охотиться на диких животных. Причиной этому, видимо, является то, что в данном возрасте они не способны решать экстраполяционные задачи, которые многократно возникают при охоте из-за отсутствия прочного предварительного опыта взаимодействия с «неисчезающими» движущимися объектами. Предположительные причины этого мы уже обсуждали. Исходя из сказанного, можно заключить, что подобные животные не могут быть пригодны для реинтродукции в природу. Судя по всему, у дикоживущих способность к экстраполяции направления движения (как проявление элементарной рассудочной деятельности) развивается по тем же принципам, что и у животных экспериментальной группы; однако в естественных условиях многообразие опыта, приобретенного в течение жизни, может быть гораздо больше. Способность применять этот опыт в решении задач, которые ставят условия обитания, несравненно повышает успешность их решения. Оценивая наши данные с той позиции, что «рассудочная деятельность не может мыслиться без «психонервных процессов», без наличия образов внешнего мира» (Беритов, 1969, с.220), мы пришли к следующему заключению, что в процессе постнатального онтогенеза способность к рассудочной деятельности развивается в два этапа. На первом этапе — в возрасте около 7 месяцев — волки решают рассудочные задачи за счет сформированной способности использовать предшествующий опыт в новой, логически схожей ситуации, т. е. в данном возрасте развивается способность «поиска» логических связей между элементами новой задачи и задач, решенных в прошлом. На втором этапе — в возрасте 1,5–2 лет у волков формируется способность прогнозировать результат своего воздействия на внешний раздражитель. Способность, развитая на данном этапе, является, видимо, наиболее сложной формой высшей нервной деятельности, формирование которой обеспечивается первым этапом. На втором этапе животное уже способно воздействовать на элементы внешней среды, изменяя их соотношение в свою пользу. Нам представляется, что оба этапа объединяет то, что решение новой задачи происходит за счет улавливания отношений, что может происходить мгновенно. Внешне, это похоже на «озарение», но только внешне. Итак, уровень, на котором происходит реорганизация и сопоставление предшествующего опыта, определяет успешность многократного решения рассудочных задач. При пассивных знаниях реорганизация и сопоставление предшествующего опыта вызывают процессы, выходящие на уровень образной психонервной деятельности, что, в свою очередь, должно вызывать повышение нервного напряжения, определяющего срывы при многократном решении рассудочных задач. В тех случаях, когда происходит оперирование знаниями, сформированными и закрепленными в течение длительного времени, не должно происходить развитие процессов, выходящих на уровень образной психонервной деятельности. Это должно обеспечить сохранение нормального уровня нервного напряжения, что, в свою очередь, определяет стабильность успешности многократного решения рассудочных задач. Исходя из всего вышесказанного, можно заключить, что условия содержания в неволе крупных хищных млекопитающих, и, в частности, волка имеют решающее значение для формирования максимального уровня (специфичного для вида) способности к рассудочной деятельности. Это говорит о том, что для реинтродукции в природу могут быть пригодны только те животные, условия содержания которых в процессе постнатального онтогенеза обеспечили максимальный уровень формирования способности к элементарной рассудочной деятельности. Резюмируя приведенный выше материал, можно сделать следующие выводы: — у волков в возрасте от 5 до 7 месяцев формируется способность применять предшествующий опыт в новой, логически схожей ситуации, что в дальнейшем облегчает им решение экстраполяционных задач; — эта способность, формирующаяся на данном этапе постнатального онтогенеза, требует постоянной тренировки, иначе ее реализация может быть значительно затруднена, что сделает животных непригодными к реинтродукции в природу; — использование прочного предшествующего опыта при решении экстраполяционных задач не вызывает процессов, выходящих на уровень образной психонервной деятельности; это обеспечивает сохранение нормального уровня нервного напряжения, что, в свою очередь, определяет высокую стабильность правильных решений при многократном предъявлении экстраполяционных задач; — при наличии определенного опыта волки способны выполнять не только элементарные «прогнозирующие» реакции при изменении соотношения раздражителей, но и прогнозировать результат целенаправленного изменения ими этого соотношения; — в процессе подготовки крупных хищных млекопитающих, и, в частности, волка к реинтродукции в природу, необходимо с ювенильного периода содержать их в таких условиях, которые обеспечат формирование максимального, характерного для данного вида уровня возможности решения рассудочных задач. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||