|
||||
|
|
РАЗДЕЛ 3. Вопросы, связанные с реинтродукциейГлава 3.1. Реинтродукция в природу выращенных в неволе крупных хищных млекопитающихМатериал и методикаМетод разрабатывался в вольерных (100 м2) и в полевых условиях, на выращенных в неволе 22 взрослых волках (Canis lupus cubanensis Ognev), разделенных на 4 группы. I группа — 2 ›, II группа — 8 › и 4 +, III группа — 2 › и 2 +, IV группа — 3 › и 1 +. У всех животных было полностью сформировано охотничье поведение, и все обладали прочным опытом. Следует отметить, что члены первой и второй групп с 2-недельного возраста росли вместе. Эксперименты в условиях неволи проводились также на 3 тиграх (P. t. altaica, 1 + и 2 ›; P. t. corbetti, 1 ›), на 3 леопардах (P. p. pardus, 1 + и 2 ›); в полевых условиях — на 20 сторожевых собаках, агрессивных по отношению к незнакомым людям, на 67 собаках, лояльных к незнакомым людям (10 групп), на 24 взрослых козах, 11 лошадях и 16 ослах. 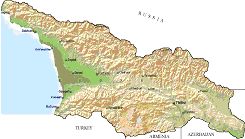 Рисунок 15. Район реинтродукции волков на Триалетском хребте. Для реинтродукции волков выбрали отрезок Мцхетско-Горийского районов Триалетского хребта (рис. 15). Выбор этих участков обусловлен тем, что вследствие принятой в тот период премиальной системы награждения за уничтожение хищников волк здесь был практически истреблен, а его экологическую нишу заняли одичавшие собаки. По нашим данным, в 1975 году на означенных участках Триалетского хребта обитало до 700 одичавших собак. В пользу этого выбора говорило и наличие пастбищ на этих участках, которые интенсивно использовались, тем самым создавая сложнейшие и поэтому наилучшие условия для апробации нашего метода. Описание выбранной для реинтродукции территорииМцхета-Горийский отрезок Триалетского хребта представляет собой крайнюю восточную часть Аджаро-Триалетской горной системы, длиной 153 километра. Ширина хребта колеблется в пределах 25–40 километров. Большая часть ширины приходится на северный склон хребта, интенсивно дренированный притоками реки Куры. На выбранном нами участке максимальная высота составляла 2249 метров. Растительный покров хребта представлен скальными ксерофитами, буком, елово-пихтовыми и сосновыми лесными массивами. В субальпийской зоне — множество лугов с хорошо развитым растительным покровом. На участке, выбранном для реинтродукции, из крупных млекопитающих обитали: около 1700 особей косули, большое количество кабанов, приблизительно 25 оленей, около 700 одичавших собак. Из мелких млекопитающих характерными являются заяц, лесная мышь и кустарниковая полевка, белка, соня, еж, белозубка, бурозубка. Условия ознакомления волков с выбранной для реинтродукции территорииДо освоения территории, выбранной для реинтродукции, у волков было сформировано охотничье поведение. За 6 месяцев до реинтродукции мы начали приучать животных к выбранной для этого территории. Ежемесячно, в течение 2 недель вывозили животных на выбранную территорию и выпускали их без применения ограничивающих движение средств. Первые 2 недели освоение территории волками происходило по нашей инициативе. Это выражалось только в выборе направления движения. Ежедневно, на протяжении 6 часов мы ходили по предварительно изученным тропам обитающих здесь зверей. Кроме этого, вместе с волками мы отправлялись к местам скопления копытных (дневные и ночные места отдыха, водопоя). В этот период животным ежедневно давали 2/3 полного мясного рациона. Через 2 недели волкам предоставляли полную самостоятельность. При этом в течение одного месяца их кормили через день, а начиная с третьего месяца, давали мясо только при их возвращении в лагерь. С этого времени перемещение животных контролировали только с помощью тропления. Для полного исключения ошибки в идентификации наших животных в порцию мяса для подкормки подкладывали куски пастэли, изготовленной на основе воска, или пищевую краску в желатиновых капсулах. Это давало возможность идентифицировать зверей по цвету экскрементов. Кроме того, анализ экскрементов производили для установления видов животных, добытых волками. За 40–50 дней до реинтродукции начинали выработку реакции избегания незнакомых людей. У животных всех видов ее вырабатывали с помощью прикрепленного к ошейнику электроразрядного стимулятора: электроды которого плотно касались шеи. Напряжение разряда составляло до 300 вольт. Реинтродукцию волков осуществляли приблизительно за неделю до начала гона. Метод выработки реакции избегания незнакомых людей и домашних животныхВолки. Во всех случаях эксперименты проводились как на сытых, так и на голодных животных. В экспериментах участвовали 36 незнакомых для животных добровольцев из представителей местного населения. Реакция избегания незнакомых людей вырабатывалась следующим образом: на участке площадью в 1 га незнакомец оставлял следы, пересекая этот участок в 2–4 направлениях. Затем на данный участок выпускали волка, который до того находился в легкой клетке с непрозрачными стенами. После изучения им участка и следов (через 3–5 мин.) появлялся незнакомец. Спустя 5 секунд после его появления, с помощью радиоуправляемого электростимулятора животное получало сильное электрическое раздражение. Такие эксперименты проводили ежедневно 4–5 раз на разных участках. После окончания каждого эксперимента волков помещали в клетку с непрозрачными стенами, чтобы они не могли видеть незнакомых людей. Во всех экспериментах добровольцы имели с собой различные предметы: рюкзак, сумку, вязанку соломы, хворост, ружье, палку; или были без всяких предметов. Максимальный возраст добровольцев — 70 лет, минимальный — 15. Описанные эксперименты проводили до выработки прочной реакции избегания незнакомых людей. Реакция считалась прочной, если волки избегали незнакомых людей без использования электрического раздражения не менее чем в 100 опытах. Кроме того, необходимым условием было отступление на дистанцию, недоступную для выстрела. Выработка реакции избегания домашних животных с помощью того же аппарата начиналась со дня освоения выбранной для реинтродукции территории. В этом эксперименте волков подводили к стаду овец или крупного рогатого скота, или к одиночным животным. При попытке приближения к ним или нападения на них хищник получал болевое электрическое раздражение. Эксперименты проводились до выработки прочной реакции избегания домашних животных, когда после 4-дневной пищевой депривации волки не подходили к домашним животным: несмотря на отсутствие электрического раздражителя. Прочность реакции испытывалась в 100 пробах. К моменту реинтродукции у всех животных выработали прочную реакцию избегания незнакомых людей и домашних животных. Наблюдения проводили в течение 4 лет после реинтродукции (в среднем 1100 часов на одну группу); наблюдались как реинтродуцированные животные, так и их потомство (первое и второе поколение). На всех реинтродуцированных животных надевали металлический ошейник с указанием его номера, моей фамилии, адреса института, а также обязательства, согласно которому человеку, добывшему данное животное, полагается денежное вознаграждение, вдвое превышающее премию, обещанную правительством за уничтожение волка (подобная премиальная система существовала до 1993 года). Это делалось с целью получения информации о возможности добывания реинтродуцированных волков. Собаки. Основной целью работы с этими животными было испытание нашего метода, как на лояльных, так и на агрессивных (в результате специальной дрессировки) по отношению к незнакомым людям собаках. Реакция избегания вырабатывалась так же, как у волков, с той лишь разницей, что у собак ее вырабатывали только относительно незнакомых людей. Кошачьи. Основной целью работы с этой группой была проверка пригодности нашего метода для других крупных млекопитающих. Ввиду нецелесообразности реинтродукции тигра и данного подвида леопарда на территорию Грузии эксперименты проводились только в условиях неволи. Всем подопытным кошачьим один раз в неделю давали живой корм (кролики, козы), а в остальное время — мясо. После полного формирования охотничьего поведения начинали выработку реакции избегания живого корма по вышеописанному методу. Домашние копытные. Выбор домашних копытных был обусловлен тем, что исторически эти животные лояльны к человеку. Все находящиеся в эксперименте животные принадлежали пастухам. Метод выработки реакции избегания незнакомых людей был таким же, как в описанных случаях. Полученные результаты и их обсуждениеСкорость выработки реакции избегания домашних животныхКак уже отмечалось, у волков вырабатывали реакцию избегания домашних животных с самого начала освоения территории, выбранной для реинтродукции. Скорость выработки реакции избегания домашних животных у волков показана в таблице 7, а в таблице 8 показана скорость выработки реакции избегания живого корма у кошачьих. 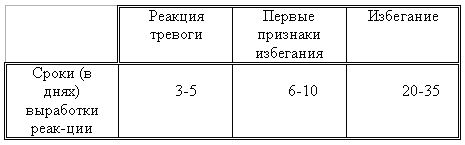 Таблица 7. Скорость выработки реакции избегания домашних животных у волков. 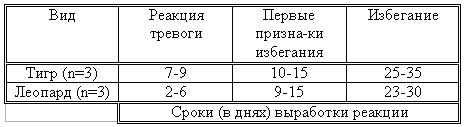 Таблица 8. Скорость выработки реакции избегания живого корма (кролики, козы) у кошачьих. Освоение выбранной для реинтродукции территорииВ первую неделю после выпуска животных на территорию, выбранную для реинтродукции, наблюдались т. н. «перебегания» между ближайшими предметами, выделяющимися своими размерами или цветом (валун, большое дерево и др.). В этот период у животных ярко выражалась реакция тревоги. После исследования указанных объектов волки в течение 1–2 минут метили их мочой или экскрементами. Затем реакция тревоги исчезала, но, как только мы вместе с волками удалялись от знакомой территории, реакция возникала снова и возобновлялось интенсивное мечение территории. Абсолютно противоположной была реакция на след зверя. В этом случае ярко выражалась интенсивная исследовательская активность, без признаков тревоги, а на экскременты — реакция Тергора. Во всех случаях волки стремились идти по следу. Надо отметить, что в течение первой недели животные шли по следу, как по его направлению, так и против него (50 % на 50 %). Со второй недели частота исследования по направлению следа возрастала и к 24-му дню достигала 90 %. В течение первых 2 недель во время совместных перемещений волки целенаправленно контролировали нашу реакцию на элементы среды и старались держаться на расстоянии визуального контакта. Находясь в поле, волки тратили большее количество энергии, чем могли восстановить из получаемой от нас пищи. Из-за систематического недоедания на 4–6 день (по группам I -5-й день, II — 4-й день, III — 6-й день, IV — 5-й день) животные стали охотиться на грызунов, вследствие чего полностью удовлетворяли свою потребность в еде. С третьей недели освоения территории волкам предоставляли возможность свободного перемещения, но, вместе с тем, продолжали систематически следовать за ними. В этот период для волков был характерен активный поиск добычи по следу. Опишем поведение волков при первом самостоятельном выходе на охоту. Первая группа (n=2). 20-й день освоения территории. Волки вышли на стадо кабанов, в котором было 3 самки и более 8 поросят 2—3-месячного возраста. Увидев кабанов, хищники сразу же напали на них, но самки и поросята разбежались в разные стороны. Волки бросились вслед за самкой, затем вернулись и стали интенсивно исследовать след. Вторая группа. (n=12) 23-й день освоения. Волки обнаружили след косули, пошли по нему и вышли на самца. Расстояние между ними составляло 50 метров. Волки и косуля одновременно заметили друг друга, в течение нескольких секунд смотрели друг на друга, после чего волки попытались напасть на нее. Косуля убежала только тогда, когда расстояние между ними сократилось до 15–20 метров. Приблизительно через 150 метров мы потеряли животных из виду и нашли только спустя 40 минут на расстоянии 400 метров от места первой встречи с косулей. Волки лежали вокруг ее скелета. Третья группа. (n=4) 25-й день освоения. Волки побежали по следу, идентифицировать который не удалось из-за сухого грунта. Приблизительно через 2 минуты я услышал звуки ломающихся ветвей, рычание волков и визг жертвы. С помощью этих звуков обнаружил волков, которые к этому моменту уже завалили подсвинка (возраст жертвы — примерно 1 год). Волки распороли добыче живот и начали есть. Четвертая группа. (n=4) 14-й день освоения. Волки случайно наткнулись на косулю, которая не заметила их. Она находилась в конечной части короткого ущелья, заканчивающегося тупиком. Волки сразу напали на косулю и убили ее на месте. * * *С третьего месяца освоения территории волки самостоятельно перемещались повсюду. Площадь территории, к концу третьего месяца освоенной волками при совместном с нами перемещении, приведена в таблице 9. Следует отметить, что первая и вторая группы были реинтродуцированы на одну и ту же территорию, с интервалом в один год. Ввиду этого во время освоения новой территории второй группой, она часто объединялись с первой.  Таблица 9. Площадь территории, освоенной волками к концу третьего месяца. В этот период волки возвращались в лагерь не более одного раза в неделю. Животные должны были привыкнуть к самостоятельному существованию, поэтому мы редко следовали за ними. Перемещение волков контролировали с помощью тропления. Работа со следом выявила, что волки любых групп избегали населенных пунктов, не приближаясь к ним ближе, чем на 2 километра. Анализ экскрементов показал, что рацион реинтродуцированных волков состоял только их диких животных. Рацион волков (по группам) к концу третьего месяца освоения территории показан в таблице 10. 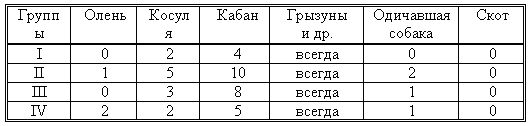 Таблица 10. Виды животных, составлявших рацион волков в течение одного месяца. Из таблицы видно, что к этому моменту освоения территории все группы добывали корм в достаточном количестве. Поэтому, начиная с четвертого месяца, животным не давали дополнительной пищи. До момента реинтродукции, т. е. до конца шестого месяца каких-либо значительных изменений с точки зрения добывания пищи или площади освоенной территории не наблюдалось. Результат выработки реакции избегания незнакомых людейЭтапы и скорость выработки реакции избегания у различных видов животных показаны в таблицах 11, 12, 13, 14.  Таблица 11. Волки (n=22). Этапы и скорость выработки реакции избегания незнакомых людей.  Таблица 12. Этапы и скорость выработки реакции избегания незнакомых людей у агрессивных собак (n=20).  Таблица 13. Этапы и скорость выработки реакции избегания незнакомых людей у лояльных собак (n=67). 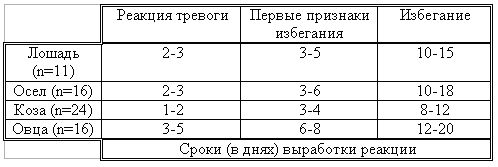 Tаблица 14. Домашние животные Результат реинтродукции волков на освоенной территории Первая группа. Группа состояла из 2 самцов, взятых в свое время из логова. Реинтродукцию осуществляли на отрезке Мцхетского района Триалетского хребта, в окрестностях горы Сатовле. К этому времени площадь освоенной волками территории составляла 150 км2. Ниже приводим материал, который касается только первого года реинтродукции. Поскольку со второго года ситуация несколько изменилась: после реинтродукции второй группы на ту же территорию, обе группы сначала объединились, а затем разделились, перераспределив территорию. В первую группу перешло четыре индивида. Далее первую группу обозначим «1+ группа». В течение первого года ежемесячно, методом анализа экскрементов (50 анализов в месяц) определяли виды животных, которыми питались волки. Анализы показали, что рацион волков состоял исключительно из диких видов. Результаты этих исследований приведены в таблице 15. 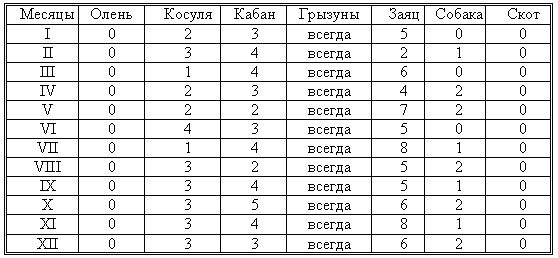 Таблица 15. Виды животных, ежемесячно входивших в состав рациона волков. Из таблицы видно, что в течение месяца волки питались только дикими видами животных. Естественно, что приведенные в таблице цифры не абсолютны, однако они полностью отражают спектр видов жертв. Особо следует подчеркнуть тот факт, что эти волки создали традицию применения определенного приема охоты, который выражался в том, что жертву выгоняли к узкому ущелью, заканчивающемуся тупиком. Это место было шириной не более 2 метров, высота стены составляла около 2,5 метров. Опишем визуально зарегистрированный случай применения этого приема. Волки погнали косулю по направлению к ущелью. На расстоянии примерно 500 метров от входа в него один волк срезал жертве путь и погнал ее в ущелье. Достигнув тупика, косуля остановилась и попыталась вернуться назад, но волки перегородили ей дорогу, атаковали и убили ее. В этом же месте мы еще раз наблюдали охоту волков. И на этот раз события разворачивались по тому же «сценарию». Тропление на данном отрезке показало, что волки часто использовали подобный прием — за последние 6 месяцев первого года мы зарегистрировали его 17 раз. Расстояние, с которого волки выгоняли жертву, колебалось от 300 до 1100 метров. Изучение останков жертв на этом месте показало, что во всех случаях это были или косули, или кабаны. В главе 1.4 мы уже обсудили способность волка к прогнозированию результата своего воздействия на внешний раздражитель и, исходя из этого, механизм формирования традиции приема охоты. Для проверки прочности реакции избегания мы с различными интервалами пытались спровоцировать нападение волков на человека и домашних животных (30 попыток). Все эти попытки были безуспешными. * * *Надо отметить, что из 12 членов второй группы в первую перешли четыре низкоранговых особи (3› и 1+). Соотношение полов в новообразованных группах было следующим: I+ группа — 5 › и 1 +; II группа — 5› и 3+. После их объединения с животными первой группы изменила статус только самка, которая, образовав пару с самцом, приобрела статус доминанта. Необходимо подчеркнуть, что до разделения объединенной (I и II) группы репродуктивный цикл начался только у доминантной самки. Интересно, что после разделения группы у перешедшей в I группу низкоранговой самки, которая в новой группе стала доминантной, через неделю начался репродуктивный цикл. После разделения между группами I+ и II началась сильная конкуренция за территорию конкуренция, которая за две недели постепенно ослабла, и к концу второй недели перераспределение индивидуальных территорий закончилось. Следует подчеркнуть, что между индивидуальными территориями групп образовалась так называемая «буферная» (нейтральная) зона, на которую, как правило, никто не выходил. Далее будет описано, как развивались события отдельно в каждой группе. I+ группа. После присоединения новых членов и перераспределения территории, владения этой группы составили приблизительно 60 км2 (рис. 16). В ее рационе за означенный период не было никаких изменений. В конце второй недели беременности самка с самцом отделились от общей группы и начали жить отдельно, но на той же территории. Уже за неделю до родов пара выбрала три пригодных для логова места. Точную дату родов установить не удалось, так как нас в это время с волками не было. Волчата родились в интервале от 15 до 29 апреля. Как оказалось впоследствии, после рождения щенков группа снова воссоединилась, и ее члены стали охотиться вместе. В рационе волков изменений не произошло. К моменту рождения волчат на их территории не было ни одной одичавшей собаки — одних убили волки, другие сами ее покинули. 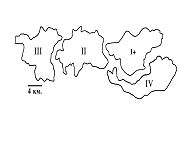 Рисунок 16. Персональные участки групп, после реинтродукции. I+ — IV — группы волков. Мы не будем касаться процесса роста и развития волчат, так как он протекал так же, как было описано в главе 2.1. Рассмотрим только процесс формирования у них реакции избегания человека и домашних животных. Наблюдения за детенышами мы стали вести после достижения ими 3-месячного возраста. Оказалось, что в стае прибавилось пять волчат. Отметим, что при первом нашем приближении к ним самка издала ранее описанный нами фыркающий лай тревоги, однако сама осталась на месте. В ответ на этот звук у волчат мгновенно появилась ярко выраженная реакция избегания. После этого случая, при любых наших попытках контакта с волчатами они активно избегали нас и внимательно наблюдали за нами с расстояния 20–30 метров. На незнакомых для родителей людей волчата реагировали по-другому. 42 раза мы пытались спровоцировать встречу детенышей с незнакомыми людьми. При первой же попытке реакция реинтродуцированных нами животных была такой же, как до реинтродукции, т. е. фыркающий лай тревоги и убегание. Поведение взрослых повторили волчата. Во всех последующих случаях детеныши самостоятельно избегали незнакомых людей. Дистанция избегания во всех случаях была достаточной. Логически схоже развивались события при встрече с домашними животными (корова, овца, коза, осел и др. — 35 случаев). И здесь, как и в случае формирования реакции избегания человека, решающее значение имело подражание реакции родителей. По достижении волчатами возраста 3 месяцев проводили анализ их экскрементов (n-57, n-40, n-40) для установления видов животных, которыми они питались. Интервал между анализами составлял 3 месяца. В последней серии анализов 27 экскрементов из 40 принадлежали волчатам и были взяты непосредственно после дефекации. Анализ показал, то все они питались только дикими животными. В период с 3 по 10 мая 1977 года у реинтродуцированных нами животных появилось второе потомство. После их рождения группа снова воссоединилась, и старшие сибсы активно участвовали в выращивании волчат. Количество новорожденных детенышей удалось установить только после достижения ими возраста одного месяца. Оказалось, что в группе прибавилось 4 волчонка. Когда им исполнилось 3 месяца, мы снова провели анализ экскрементов со стандартным интервалом (n-73, n-85, n-70). Анализ показал, что все члены группы продолжали питаться только дикими животными. По остаткам пищи определили интервал между удачными охотами взрослых. Он составил 3–4 дня. В этом интервале все волки интенсивно охотились на грызунов. В этот же период проверили прочность реакции избегания у волков относительно человека и возможность формирования этой реакции у новой генерации. Проведенные 60 экспериментов показали, что у всех реинтродуцированных нами животных реакция избегания сохранилась без всяких изменений. У старших сибсов реакция избегания относительно нас и незнакомых людей была четко дифференцирована. Нас они подпускали на расстояние 10–25 метров, а незнакомых людей — не менее, чем на недосягаемое для выстрела расстояние. У прибылых указанная реакция вырабатывалась за счет одноразового обучения, т. е. так же, как в свое время у их старших сибсов. И в этом случае хорошо была выражена дифференциация интенсивности реакции избегания относительно нас и незнакомых людей. В возрасте 7–8 месяцев погибли два щенка. Судя по внешним симптомам, причиной гибели была т. н. «собачья чума». Следует отметить, что, когда молодым животным исполнилось 8 месяцев, в группе возникли сильные агрессивные взаимодействия, в которых не принимали участия молодые особи. В результате возникшего конфликта из группы «выдворили» троих низкоранговых индивидов, из которых один — старший сибс (возраст 1 год 8 месяцев) и два — реинтродуцированные нами волка. В течение одного месяца не удавалось установить местонахождение этих волков, но затем их обнаружили между территориями I+ и II групп, в т. н. «буферной зоне». Надо отметить, что они часто пытались вернуться в свою группу, но оставшиеся в ней волки, включая молодых, встречали их сильной агрессивной реакцией. Изгнанные индивиды пытались также проникнуть на территорию соседней стаи, что также закончилось конфликтом. Именно во время конфликта с соседней стаей погибли два индивида, судьба третьего осталась неизвестной. II группа. После перехода четырех индивидов в первую группу и перераспределения территории данная группа заняла площадь в 65 км2. За этот период изменений в рационе группы не было. На третьей неделе беременности самка, а вместе с ней и самец отделились от стаи, но остались на той же территории. За неделю до родов мы обнаружили только одно логово. Установить точную дату родов волчицы не удалось из-за нашего отсутствия в этот период. Волчата родились в интервале между 15 апреля и 2 мая 1976 года. К этому периоду изменений в рационе волков не было. К моменту рождения волчат на территории стаи не было уже ни одной одичавшей собаки. Так же, как и в случае I группы, часть собак была убита волками, другая часть покинула данную территорию. Наблюдения за волчатами начали по достижении ими 3-месячного возраста. Оказалось, что к стае прибавилось 6 волчат. Трое погибли в возрасте 4 месяцев в результате заболевания «собачьей чумой». При первой попытке установления контакта с волчатами события развивались так же, как и в группе I+. В частности, самка издала описанный нами ранее лай тревоги, но сама осталась на месте. В ответ на этот лай у щенков появилась ярко выраженная реакция избегания. После этого случая, при каждой попытке контакта щенки активно избегали нас и наблюдали за нами с расстояния 15–30 метров. По-другому развивалась реакция волчат относительно незнакомых для родителей людей. Мы попытались спровоцировать встречу незнакомых людей со щенятами 38 раз. При первой же попытке у реинтродуцированных животных была реакция такой же интенсивности, как и перед реинтродукцией, т. е. фыркающий лай тревоги и убегание. Во всех последующих случаях волчата самостоятельно избегали людей. Дистанция избегания каждый раз была достаточной. Аналогично развивались события и при предъявлении домашних животных (30 случаев). И здесь, как и при формировании реакции избегания относительно человека, решающее значение имела реакция родителей. Анализ экскрементов, проведенный со стандартным интервалом (n-67, n-83, n-53) показал, что все волки питались только дикими видами. В период между 3 и 13 мая 1978 года у реинтродуцированных нами животных появилась вторая генерация. После рождения второй генерации группа заново воссоединилась 3, и старшие сибсы стали активно участвовать в воспитании детенышей. Определить число новорожденных удалось только после того, как им исполнился 1 месяц. Оказалось, что родились 4 волчонка. Когда им исполнилось 3 месяца, провели повторный анализ экскрементов со стандартным интервалом (n-88, n-78, n-73). Анализ показал, что все члены группы питались только дикими животными. В этот же период у волков проверили прочность реакции избегания человека и возможность ее формирования у нового поколения. Было проведено 35 экспериментов, которые показали, что у всех реинтродуцированных нами животных реакция избегания незнакомых людей была такой же, как и в момент реинтродукции. Родившиеся же уже на воле, избегали нас на расстоянии 10–25 метров, а незнакомых людей — на достаточно безопасное расстояние. У новой генерации эта реакция сформировалась в результате одноразового опосредованного обучения, т. е. так же, как и у их сибсов. Когда волчатам исполнилось 8 месяцев, в группе между старшими волками стали развиваться сильные агрессивные взаимодействия, в которых молодые не принимали участия. В результате возникшего конфликта из группы изгнали четырех реинтродуцированных нами низкоранговых индивидов. Последние поселились на смежной свободной территории, площадь которой через месяц составила примерно 40 км2. Следует отметить, что покинувшие группу индивиды в течение 2 недель пытались вернуться на прежнюю территорию, но каждый раз наталкивались на сильное агрессивное сопротивление. В процессе мечения «изгнанниками» границ новой, освоенной территории, сформировалась «буферная зона» вблизи с границами I и II групп. III группа. Реинтродукция осуществлена 10 января 1978 года вблизи деревни Зирта. Спустя месяц территория группы составили приблизительно 70 км2. Следует отметить, что в течение первой недели восточная граница территории этой группы непосредственно соприкасалась с границей II группы, из-за чего между членами двух групп постоянно отмечали сильные агрессивные взаимодействия. К концу первого месяца реинтродукции расстояние между границами групп составило от 1,5 до 3 км. С возникновением нейтральной зоны агрессивные взаимоотношения прекратились. В течение первого месяца в рационе группы изменений не было. В конце второй недели беременности доминантная самка и доминантный самец отделились от остальной группы и начали жить отдельно, оставаясь, однако, на той же территории. Уже за 2 недели до родов волки выбрали два пригодных для логова места. Волчата родились между 27 и 28 апреля 1978 года. Через 5 дней после их рождения группа снова воссоединилась, и ее члены стали охотиться вместе. После рождения первой генерации рацион волков остался без изменений. Так же как и в описанных выше случаях, к моменту рождения волчат на территории стаи уже не было ни одной одичавшей собаки — часть уничтожили волки, часть ушла за пределы данной территории. Наблюдать за волчатами мы начали по достижении ими 3-месячного возраста. Оказалось, что в стае прибавилось 6 волчат. Следует отметить, что при первой попытке контакта с детенышами, как и в случае с предыдущими группами, волчица издала фыркающий лай тревоги, но осталась на месте. В ответ на эти звуки у волчат появилась ярко выраженная реакция избегания. После этого случая щенки активно избегали нас и интенсивно наблюдали за нами с расстояния около 20–30 метров. Иначе развивалась реакция волчат на незнакомых для родителей людей. 42 раза мы пытались спровоцировать встречу щенят с незнакомыми людьми. При первой же попытке у реинтродуцированных нами животных проявлялась такая же реакция, как и перед реинтродукцией, т. е. фыркающий лай тревоги и убегание. Поведение родителей вслед за ними повторили и детеныши. Во всех последующих случаях волчата самостоятельно избегали незнакомых людей. Дистанция избегания всегда была достаточной. Логически схоже развивались события и при предъявлении волкам домашних животных (35 случаев). И здесь, как и при формировании реакции избегания человека у волчат решающее значение имела реакция родителей и подражание ей… После достижения волчатами 3-месячного возраста был проведен анализ экскрементов со стандартным интервалом (n-69, n-68, n-50). Анализ показал, что они питались только дикими животными. В период с 3 по 10 мая 1979 года у реинтродуцированных нами животных этой группы также появилась вторая генерация. После родов группа воссоединилась, и старшие сибсы принимали активное участие в воспитании волчат. Количество новорожденных удалось установить только спустя месяц после их рождения. Оказалось, что родилось четверо волчат. После достижения ими 3-месячного возраста были проведены анализы экскрементов со стандартным интервалом (n-43, n-70, n-57). Анализ показал, что все волки, как и прежде, питались дикими животными. В этот же период проверили прочность реакции избегания и возможность ее формирования у нового поколения. Было проведено 60 экспериментов, которые показали, что у всех реинтродуцированных животных реакция избегания сохранилась без изменений. У старших сибсов реакции на нас и на незнакомых людей были четко дифференцированы, как и во всех вышеуказанных случаях. Они избегали нас, начиная с расстояния 10–25 метров, незнакомых людей — с более безопасного расстояния. У нового поколения эта реакция выработалась в результате опосредованного обучения — в данном случае, в основном, на основе подражания реакциям старших сибсов. И в этом случае хорошо выражалась дифференциация реакции избегания нас и незнакомых людей. В период между 7 и 8 месяцем погибли два щенка. Судя по внешним симптомам, причиной смерти, опять таки, была т. н. «собачья чума». Следует отметить, что, когда молодым волкам исполнилось 8 месяцев, в группе (как и в вышеописанных случаях) возникли ярко выраженные агрессивные взаимодействия, в которых не участвовали молодые особи. В результате конфликта из группы были изгнаны 3 особи, из которых один — старший сибс в возрасте около 20 месяцев, и два — реинтродуцированных нами индивида. Через некоторое время изгнанных волков обнаружили в т. н. «буферных зонах», находящихся между территориями II и III групп. За 1,5 месяца нахождения здесь эти животные систематически пытались вернуться в прежнюю группу, но ее члены, включая молодых, встречали их сильной агрессией. Кроме того, они пытались вторгнуться и на территорию соседней группы, что также завершалось конфликтом. В конце концов «изгнанники» заняли свободную территорию северо-восточнее III группы. В течение 3 недель между территориями образовалась нейтральная зона шириной около 1–2,5 км. IV группа. Реинтродукция осуществлена 25 января 1979 года вблизи урочища Нахширгора. Через месяц после реинтродукции, территория группы составляла приблизительно 65 км2. Северная граница территории этой группы непосредственно примыкала к границам групп I+ и II, вследствие чего между членами IV, I+ и II групп развивались сильные агрессивные взаимодействия. К концу первого месяца реинтродукции расстояние между границами этих двух групп составило от 1-го до 3-х км. С возникновением нейтральной зоны агрессивные взаимодействия прекратились. В течение первого месяца рацион группы оставался без изменений. В конце второй недели беременности доминантный самец и доминантная самка отделились от остальной группы и начали жить отдельно, но на той же территории. Уже за 2 недели до родов волки выбрали два места, пригодных для логова. Точную дату рождения волчат установить не удалось из-за нашего отсутствия в этот период. По-видимому, волчата родились между 25 апреля и 5 мая 1979 года. После их рождения группа заново воссоединилась. Рацион волков не изменился. К моменту рождения волчат на территории стаи, как и во всех остальных случаях, не было ни одной одичавшей собаки. Наблюдать за волчатами начали по достижении ими 3-месячного возраста. Оказалось, что в группе прибавилось 6 волчат. Следует отметить, что при первой попытке контакта с детенышами события развивались так же, как в вышеописанных случаях. Затем щенки активно избегали нас на дистанцию около 10–20 метров. Как и в остальных случаях, заметна была четкая дифференциация между реакциями на нас и на незнакомых родителям людей. 30 раз мы пытались спровоцировать встречу щенят с незнакомыми людьми. При первой же попытке у реинтродуцированных нами животных проявлялась такая же реакция, как и перед реинтродукцией. Поведение родителей вслед за ними повторили и детеныши. Во всех последующих случаях волчата самостоятельно избегали незнакомых людей. Дистанция избегания всегда была достаточной. Логически схоже развивались события и при предъявлении волкам домашних животных (47 случаев). И здесь, как и при формировании реакции избегания человека у волчат решающее значение имела реакция родителей и старших сибсов. После достижения волчатами 3-месячного возраста был проведен стандартный анализ экскрементов (n-35, n-62, n-54). Анализ показал, что в их рацион не входили домашние животные. Через год, в период с 15 по 25 апреля 1980 года у реинтродуцированных нами животных IV группы появилось второе поколение. Старшие сибсы активно включились в воспитание детенышей. Количество новорожденных удалось установить только спустя 1,5 месяца. Оказалось, что родилось 5 волчат. После того, как им исполнилось 3 месяца, провели серию стандартных анализов экскрементов (n-65, n-82, n-40), который показал, что и все эти волки также питались только дикими животными. В этот же период, как и всегда, проверили прочность реакции избегания человека и возможность ее формирования у нового поколения. Было проведено 30 экспериментов, которые показали, что у всех реинтродуцированных животных реакция избегания сохранилась без изменений. У старших сибсов реакция на нас и на незнакомых людей были четко дифференцированы, как и во всех вышеуказанных случаях. Они избегали нас на расстоянии 10–25 метров, незнакомых людей — на достаточное расстояние. У нового поколения эта реакция выработалась по такому же принципу, что и во всех предыдущих случаях — в результате одноразового опосредованного обучения. Как видно из представленного материала, скорость выработки реакции избегания, ее дифференцированность и прочность во всех группах были одинаковыми. В связи с тем, что во всех реинтродуцированных нами группах явления развивались одинаково, можно даже сказать, стереотипно, на данном этапе наблюдений мы сочли работу завершенной. Надо отметить, что кратковременные наблюдения за отделившимися группами и их потомством (около 50 часов на каждую группу) показали, что у всех животных, включая потомство, полностью сохранена реакция избегания людей и домашних животных. Анализ экскрементов, проведенный два раза в течение года (всего 115 анализов), показал отсутствие домашних животных в рационе волков. Проводимые ежегодно опросы (по 1987 год включительно) местного населения и пастухов не выявили фактов хищничества волка в отношении домашних животных. Отмечены были три случая хищничества медведя. Ни одного случая возврата ошейников реинтродуцированных нами волков не было. Полученный материал однозначно указывает на то, что все 22 реинтродуцированных нами волка в течение всего периода наблюдений активно избегали незнакомых людей и не нападали на домашних животных. Более того — они целенаправленно обучали свое потомство тем навыкам, которым в свое время мы обучили их. Необходимо подчеркнуть и то, что старшие сибсы так же целенаправленно обучают младших тем навыкам, которым они сами обучились у родителей. Таким образом, все 35 волков выживших из 40 прибылых, активно избегали незнакомых людей и не нападали на домашних животных. Надо специально отметить, что одним из решающих условий успеха реинтродукции волков было участие представителей местного населения в этом процессе, что полностью сняло проблему их антагонизма к реинтродуцируемому виду. Это произошло потому, что реакцию избегания людей и домашних животных мы вырабатывали при их непосредственном участии в результате чего люди убеждались в том, что волк может стать неопасным для них самих и их собственности. Для успешной охоты, помимо формирования ее элементов в процессе постнатального онтогенеза, животным необходимо детальное знание территории, т. е. формирование т. н. «мысленной карты», которая обеспечивает свободное ориентирование на ней — животному необходимо знать, как и где обнаружить добычу, затаиться или просто перемещаться в нужном направлении. Разработанный нами метод дает возможность решить и эту задачу. С учетом полученного нами материала и специфики высшей нервной деятельности, этот метод может быть применен к любому виду крупных наземных млекопитающих. Таким образом, становится возможным решение проблемы восстановления исчезающих или исчезнувших в природе видов путем реинтродукции в естественную среду обитания выращенных в неволе индивидов. Глава 3.2. Реакция диких копытных на хищных: — инстинкт или традиция?Знание поведенческих взаимоотношений хищника — жертвы приобретает особое значение тогда, когда происходит восстановление находящихся на грани исчезновения или исчезнувших в природе видов, независимо от того, хищник это или копытное. Когда речь идет о защитной реакции копытного по отношению к хищнику, подразумевается тот поведенческий комплекс, который выражается в убегании, вокализации и в поведении, непосредственно связанном с охраной себя или потомства. Вальтер (Walter, 1970) показал, что «дистанция бегства» газели Томпсона — Gasella tomsoni (подразумевается то минимальное расстояние между хищником и потенциальной жертвой, при сокращении которого она убегает от хищника) зависит от пола, возраста и ранга индивида. Интересно, что у взрослых самцов эта дистанция намного меньше, чем у самок. У самок же — приблизительно такая же, как у молодых самцов. У самцов, имеющих пары, дистанция бегства меньше, чем у самцов, не имеющих таковых, а у последних, в свою очередь, меньше, чем у не территориальных самцов, у которых дистанция убегания больше, чем у самок. Автор считает, что внутрипопуляционная дифференциация дистанции бегства является, в первую очередь, средством поддержания социальной структуры популяции и только потом — адаптацией, позволяющей избежать нападения хищника. Представленная здесь интерпретация автора интересна сама по себе и полностью приемлема. Но мы хотим обсудить те стороны проблемы, которых автор не коснулся. Исходя из цитированного материала, можно сделать еще один вывод: меньшая «дистанция бегства» доминантных индивидов обуславливает еще и большую вероятность гибели их (производителей) от хищника. Это, в свою очередь, обеспечивает смену доминантов и способствует сохранению их оптимального возраста (производителей), как это уже было показано раньше (Бадридзе и соавт., 1992). Дистанция бегства, как одна из пороговых форм защитного поведения, в свое время была описана Баскиным (1976). В его работе описана дистанция бегства некоторых видов копытных от человека. Как выясняется, она различна у разных видов. Из приведенной работы можно сделать вывод — дистанция бегства зависит от потенциальной скорости передвижения (или бега) вида и характерного для него психологического типа. Особого внимания заслуживает работа Шаллера (Schaller,1972), в которой автор показал, что дистанция бегства зебры от хищника зависит от вида самого хищника. Так, например, эта дистанция при убегании от пятнистой гиены равна 9 м., от гиеновой собаки — 18 м., льва — 45 м., а если в стаде находятся жеребята, то и 90 м. Следовательно, дистанция бегства от хищника прямо пропорциональна потенциальной скорости последнего; в то же время учитываются скоростные возможности находящихся в стаде жеребят. Несмотря на то, что приведенными трудами ограничиваются данные по рассматриваемой теме, они полностью отражают сущность указанного поведенческого феномена. Однако они, к сожалению, не отвечают на поставленный нами вопрос: является ли защитная реакция копытных инстинктом на хищника или приобретенной формой поведения, ставшей традицией? Поэтому нашей целью было изучение защитного поведения (поведенческого иммунитета) копытных по отношению к хищнику, природы данного поведения и механизма его формирования. Изучение этого вопроса поможет определить оптимальный темп реинтродукции хищника. Материал и методикаНаблюдения и эксперименты проводили в природных условиях на 860 кавказских благородных оленях (Cervus elaphus marsal) с оленятами и на 300 дагестанских турах (Capra cilindricornis) с козлятами. Изучали реакцию диких копытных на человека и на хищных. Для изучения защитной реакции на человека наблюдения и эксперименты проводили на территории Грузии, в регионах, подвергшихся наибольшему прессу браконьерства (Триалетский хребет — Мцхетский, Горийский, Тетрицкаройский районы, Боржомский заповедник, Кахетинская часть Большого Кавказа, включая Лагодехский заповедник). Кроме этого, специальные наблюдения проводили в Лагодехском заповеднике в период интенсивного браконьерства (1992–1994 годы). Для определения уровня браконьерства систематически считали услышанные выстрелы и определяли направление, откуда они доносились. одним фактом браконьерства считали случай, когда выстрелы раздавались из одного и того же места, независимо от их количества. Для каждых 7 дней вычисляли коэффициент уровня браконьерства k=n/7, где n-число ежедневных случаев браконьерства. Исходя из полученного коэффициента, устанавливали влияние уровня браконьерства на дистанцию бегства копытных. Изучение реакции диких копытных на человека велось двумя методами: I — приближались открыто; II — приближались скрытно на возможно минимальную дистанцию, после чего появлялись перед животными. В обоих случаях описывали поведенческую реакцию копытных и их детенышей, вокализацию и дистанцию бегства. При изучении реакции копытных на хищников наблюдения и эксперименты проводили по отдельности, в регионах, где на протяжении нескольких лет не было волков (территории Мцхетского и Горийского районов на отрезке Триалетского хребта), а также в регионах, где волки обитали постоянно (Боржомский и Лагодехский районы). В Триалети наблюдения вели за дистанцией бегства копытных от реинтродуцированных нами волков. В других же районах — подкрадываясь на 200–300 метров к группе копытных, выпускали выращенного в неволе волка с намордником (из 53 экспериментов в 20 случаях волк не умел охотиться) и наблюдали за их реакциями. Полученные результаты и их обсуждениеПрежде чем изложить материалы, полученные в результате наблюдения за животными, целесообразно привести таблицу коэффициентов, отражающих уровень браконьерства в изученных нами регионах (таблица 16). 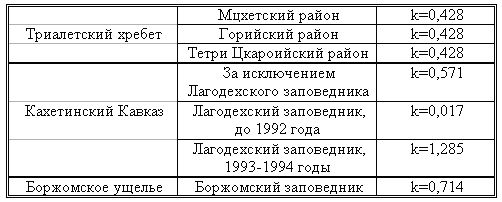 Таблица 16. Коэффициент уровня браконьерства в различных регионах Грузии. Изучение дистанции бегства от нас диких копытных показало, что ее показатель находится в прямой зависимости от коэффициента (к) уровня браконьерства. Так, например, дистанция бегства от нас была минимальной в тех районах, где к=0,017 (кавказский олень — 30–50 м), а там, где коэффициент браконьерства был максимальным (к=1,285) соответственно увеличивалась и дистанция бегства (кавказский олень — 300–500). Детально эти данные приведены в таблице 17. 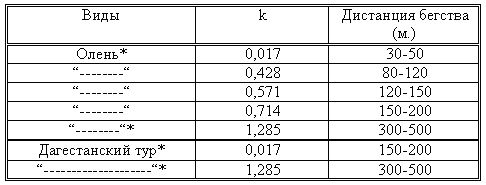 Таблица 17. Влияние уровня браконьерства на дистанцию бегства копытных от человека. * — одни и те же популяции. Данные об изменении уровня браконьерства в одном и том же регионе получены нами только в Лагодехском заповеднике. Однако и их достаточно, чтобы было очевидным влияние этого фактора на дистанцию бегства представителей одной и той же популяции (рис. 17). Полученный материал однозначно указывает на то, что дистанция бегства не является врожденной. Она может изменяться пропорционально изменению пресса браконьерства на копытных. Исходя из этого, об уровне браконьерства можно судить по дистанции бегства копытных от человека. По нашему мнению, это имеет большое практическое значение для оперативного выявления ситуации в том или ином районе. 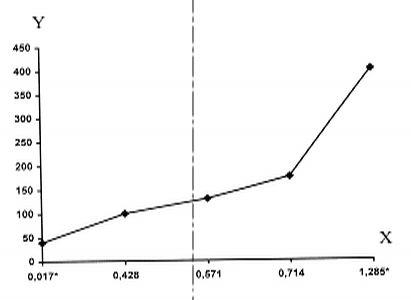 Рисунок 17. Зависимость дистанции бегства Кавказского благородного оленя от уровня браконьерства. X — коэффициент браконьерства; Y — средняя дистанция бегства. * — одни и те же популяции. Следует отметить тот факт, что дистанция бегства существующих в группе молодых индивидов и взрослых животных одинакова. Мы подчеркиваем, существующих в группе, т. к. дистанция бегства молодых может измениться после того, как они выйдут из-под опеки родителей и начнут самостоятельную жизнь. Полученный нами материал говорит и о том, что дистанция бегства взрослых индивидов меняется в зависимости от индивидуального (или группового) опыта. Потомству же опыт передается путем опосредованного обучения. Исходя из этого, следует подчеркнуть, что дистанция бегства индивида не является врожденной и передается следующему поколению за счет врожденной способности к подражанию. Последнее же — один из значительных механизмов обучения. Эксперименты, проведенные при подкрадывании к диким копытным на возможно минимальную дистанцию, показали, что самки оленей и туров с детенышами при внезапном появлении человека на близком расстоянии издают специфические звуки (олени — резкий лающий звук, туры — свист). Вслед за этим убегают взрослые особи; детеныши же, как правило, убегают сразу при звуке тревоги. В некоторых случаях (n=27) детеныши убегали только вместе с родителями. При повторном подкрадывании к этим индивидам детеныши убегали не вместе с родителями, а на фоне звуков тревоги, издаваемых родителями. Описанные факты указывают на то, что реакция убегания детенышей формируется за счет одноразового обучения. Для этой формы обучения решающее значение имеет врожденная реакция на издаваемый родителем звук тревоги и сопряженная с этим звуком поведенческая реакция того же родителя. Следует подчеркнуть и то, что этот звук является для детеныша безусловным раздражителем. Именно сопряжение этого раздражителя с появлением человека обуславливает формирование защитного поведения детенышей за счет одноразового обучения. При сопоставлении материала, полученного нами на волках и копытных, стало очевидным, что и у хищных, и у копытных в процессе постнатального онтогенеза описанная поведенческая реакция должна формироваться по одному и тому же принципу. Можно предположить, что поведенческая реакция родителей формирует традицию конкретной формы защитного поведения потомства и ее передачу от поколения к поколению путем опосредованного обучения. На основе вышесказанного можно сделать вывод, что ситуационное выражение и интенсивность защитной формы в зависимости от поведения не является врожденной и поэтому может меняться в зависимости от состояния окружающей среды. Этим обеспечивается формирование оптимальной формы поведения и ее интенсивности, и тем самым адаптация к условиям окружающей среды. Наблюдения за охотой подготовленных нами волков показали, что с момента реинтродукции в течение 6 месяцев дистанция бегства от них диких копытных возросла приблизительно в 6 раз. Изменение дистанции бегства показано на графике 18. 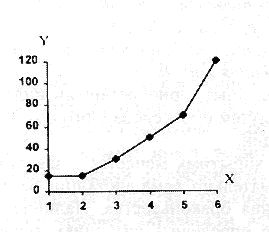 Рисунок 18. Изменение дистанции бегства косуль от реинтродуцированных волков. X — месяцы со дня реинтродукции; Y — дистанция бегства в метрах. 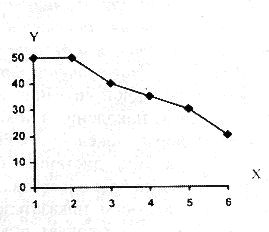 Рисунок 19. Изменение процента успешной охоты реинтродуцированных волков. X — месяцы со дня реинтродукции; Y — процент успешной охоты. Как видно из приведенного материала, между дистанцией бегства от хищника и успешностью охоты последнего существует отрицательная корреляция (рис. 19). Для формирования такой корреляции, по-видимому, большое значение имеет опыт, приобретенный «спасшейся» жертвой. Этот опыт использует не только она сама, но и передает его другим (включая потомство) путем опосредованного обучения. Интересно и то, что дистанция бегства копытных увеличивается и в том случае, когда индивиды наблюдают смерть своих собратьев. Поэтому обучение возможно не только за счет собственного опыта или при наблюдении за завершенной поведенческой реакцией других особей, но и тогда, когда другой индивид успевает осуществить лишь небольшой фрагмент защитного поведения и оно кончается неудачей. Исходя из этого, одноразовое сочетание внешних признаков врага (хищник, человек) и защитного поведения или его фрагмента у собрата достаточно для формирования адекватной защитной реакции животного — «наблюдателя». Сразу же после восстановления у копытных защитной реакции по отношению к волку в вышеуказанных регионах провели эксперименты с целью изучения их реакции на хищников, не имеющих опыта охоты. Эксперименты показали, что во всех 20 случаях у копытных развивалась такая же четко выраженная защитная реакция, как и на особей, имеющих опыт охоты. Это еще раз указывает на решающее значение видовых признаков хищника для формирования у копытных оборонительной реакции. Эксперименты, проведенные в регионах постоянного обитания волка, показали, что реакция диких копытных на волков четко дифференцирована. В частности, на имеющих опыт охоты хищников у копытных имеется ярко выраженная защитная реакция, тогда как они индифферентны по отношению к тем особям, которые не умеют охотится, т. е. не воспринимают копытных как потенциальную жертву. Такие индивиды могли находиться в непосредственной близости к оленям. Приведенный материал перекликается с работой Мичурина (1970), где он отмечает, что часто видел волков, которые разгуливали среди группы оленей. Расстояние между ними менялось от 30 до 100 метров. В другом случае волк сидел среди большой группы оленей на холме. Приведенный нами материал указывает на то, что при взаимодействии с хищником копытные приобретают опыт, который позволяет им различать не только его видоспецифичные черты, но и специфику поведения этого вида и, исходя из этого, и его «цель». Из всего вышесказанного можно сделать следующий вывод: реакция диких копытных на человека и хищников не является врожденной. Форма этой реакции и ее интенсивность могут изменяться соответственно уровню пресса на копытных. Приобретение опыта (обучение), может происходить с одного раза, не только во время непосредственно полученного «урока», но и методом наблюдения за реакцией других индивидов (опосредованное обучение). Для опосредованного обучения достаточно восприятия фрагмента реакции другого индивида, что полностью исключает необходимость приобретения собственного опыта, тем самым, значительно ускоряет процесс обучения и делает его значительно безопасным и, следовательно, имеет большое значение для выживания индивида. В случае стабильной ситуации в ареале, опыт, приобретенный индивидом, передается из поколения в поколение путем опосредованного обучения и приобретает форму поведенческой традиции. Как было показано, дистанция бегства копытных меняется в зависимости от уровня пресса браконьерства. Следовательно, сама дистанция бегства является безошибочным показателем уровня браконьерства. Наличие подобной зависимости позволяет оперативно определить ситуацию, существующую в том или ином регионе, что имеет важной практическое значение для контроля за уровнем браконьерства. Если интенсивность описанной защитной реакции не является врожденной, то она может угаснуть или ослабнуть в том случае, если в ареале копытного, по разным причинам, больше нет хищника. Наглядным примером этого служит дистанция бегства косули от реинтродуцированных нами волков. Как оказалось, эта поведенческая форма и ее интенсивность восстановимы, но в масштабе популяции для ее восстановления необходимо длительное время. Исходя из этого, в процессе восстановления вида хищника очень важно, какое количество животных реинтродуцируется одновременно. Реинтродукция одновременно большого числа хищных до восстановления защитного поведения копытных может отрицательно сказаться на их популяцию. Анализ приведенного материала позволяет сделать следующие выводы: 1. форма и интенсивность защитной реакции копытных по отношению к врагу (человеку или хищнику) подвержены сильной модификации в онтогенезе, вследствие чего возможны их изменения во времени в соответствии с существующей ситуацией; 2. в конкретной ситуации форма и интенсивность защитного поведения по отношению к врагу передается опосредованным путем из поколения в поколение; 3. у копытных, в случае отсутствия в их ареале хищников, через одно поколение гаснет оборонительная реакция по отношению к ним; 4. во избежание отрицательного воздействия на численность копытных, реинтродукция (судя по всему, и транслокация) восстанавливаемого вида хищников должна производиться небольшими группами и со значительными интервалами. Естественно, что интервал должен определяться, исходя из поведенческой и репродуктивной специфики копытных; 5. пресс браконьерства по отношению к копытным можно определять по дистанции их бегства от человека. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||