|
||||
|
|
Глава двенадцатая. Беспозвоночные и рыбы Основы истории развития животных Геологическая история развития растений дает хорошую основу для изучения истории животных благодаря тесным био" мическим связям этих двух групп организмов: растения ом спечивают животных пищей и свободным кислородом, а животные поставляют необходимый растениям углекислый га Это взаимоотношение позволяет считать, что обе группы oрганизмов развивались более или менее параллельно. В предыдущей главе мы прочли, что эволюция некоторых животнш происходила под влиянием определенных видов растений и что всем этим управляли процессы, происходящие в недрах и на поверхности Земли. Наше исследование истории развития животных, как и истории растений, основывается на данных изучения ископаемых остатков. Но летопись ископаемых остатков животных имея много пробелов, и пока не будут найдены руководящие ископаемые, мы вынуждены заполнять эти пробелы умозрительным! рассуждениями. По мере того как мы обнаруживаем и изучает все новые ископаемые остатки, некоторые из построений предыдущих лет подтверждаются и становятся доказанными истинами, другие оказываются неверными и заменяются положениями, более соответствующими фактам. Развитие растительного и животного мира Как мы уже говорили, между животными и растениями существует коренное различие, возникшее с самых первых шага развития жизни на Земле. Это различие заключается в способе питания. Растение нуждается в основном в углекислом газе и солнечном свете, с помощью которых оно путем фотосинтеза вырабатывает органические вещества, служащие ему пищей и образующие ткани его тела. Животные, как одноклеточные, так и многоклеточные, используют в пищу ткани других организмов: или растений, или вторично других животных. Кроме того, они дышат кислородом, а этот источник энергии также поставляется в атмосферу и гидросферу растениями. Без пищ и кислорода животные не могли бы существовать. В таком случае они, очевидно, не могли возникнуть до тех пор, пока не появились растения и не начали в процессе фотосинтеза выделять свободный кислород. Как было показано в таблице 2, мы располагаем данными летописи ископаемых, свидетельствующими о том, что в начале последнего миллиарда лет истории Земли (до фанерозоя) и растения, и животные жили в море, взаимно дополняя процессы жюнедеятельности друг друга. Растения высвобождали кислород из углекислого газа, а животные дышали этим свободным кислородом, окисляли растительную пищу и создавали углекислый газ, который снова использовался растениями. Таково биохимическое взаимодействие растений и животных. Однако, хотя это взаимодействие продолжалось и позднее, животные начинали приобретать все более и более отличный от растений облик. Растениям, жившим сначала в мелководных морях, а затем на суше, не нужно было перемещаться в поисках пищи, они могли просто поглощать ее, оставаясь на месте. На суше им для этого требовались только листья, каждый из которых освещался солнцем, корневая система для поглощения водных растворов из грунта и соединительное устройство, служащее для поддержки листьев и подачи к ним растворов. Именно такими приспособлениями обладает большинство растений. Животным же, за исключением некоторых, наиболее простых, прикрепленных к морскому дну, приходилось двигаться в поисках пищи, которой им служат растения и другие животные. Животные - это охотники, и с самых первых шагов эволюции они приобрели способность к передвижению. Наиболее древние животные, жившие в море, изменялись различными способами. Путь самого успешного развития представляет для нас наибольший интерес, так как в ходе его были заложены основы строения человеческого тела. Это развитие состояло в образовании: во-первых, прочного жесткого скелета, к которому прикреплялись мускулы; во-вторых, хвоста, который использовался как рулевое весло; в-третьих, системы конечностей (равного числа правых и левых), которые могут двигаться с помощью мускулов, чтобы уравновешивать тело в воде и затем чтобы перемещать его в толще воды; в-четвертых, глаз, расположенных в передней части тела для того, чтобы видеть добычу и препятствия, которые нужно избегать; в-пятых, рта не только для того, чтобы поглощать пищу, но также и воду; в-шестых, центрального управляющего устройства - мозга, соединенного с мускулами нервной системой. Если мы добавим к этому списку седьмой пункт - удлиненную, обтекаемую форму тела, служащую для уменьшения трения при движении в толще воды, то становится понятно, что получившийся в результате организм в основном подобен рыбе. Конечно, не все эти усовершенствования появились сразу. Они развивались в течение десятков, если не сотен миллионов лет. Ни одно из них не было чем-то совершенно новым. Все они представляли собой дальнейшее развитие или видоизменение уже существовавших свойств. Рыбообразная форма тела была наиболее удачной для жизни в море. Позднее эта же конструкция послужила основой для формирования тела наземных животных. Условия обитания организмов в море Нетрудно представить себе, что древнейшие животные должны были развиваться во множество видов, потому что океан далеко не однообразен. В нем можно выделить множество различных видов среды, в которых происходит (путем естественного отбора) формирование организмов, характеризующихся многообразием форм и специализаций. Миллиард лет назад в океане, как и сейчас, животные могли плавать на поверхности воды или вблизи нее, могли населять холодное темное дно и толщу воды между дном и поверхностью. На мелководье вблизи берегов, как и сейчас, условия жизни различались в зависимости от температуры, вида коренных пород или рыхлых отложений, слагающих дно, интенсивности перемешивания воды волнами или течениями. Каждое даже самое небольшое отличие условий среды оказывало влияние на организмы путем естественного отбора. Морские животные приобретали специализацию в соответствии с условиями жизни. Некоторые из них были растительноядными, поедавшими морские растения, другие - плотоядными, питавшимися другими животными. Часть из них представляла собой так называемых "мусорщиков" и поедала трупы животных. Другие были хищниками, пожиравшими живую добычу, Несомненно, что уже тогда, в эти древние времена, мелководные моря были оживленными местами; их многочисленные обитатели занимались поисками пищи, спасались от хищников, поедали других животных или сами становились добычей.
Фото 29. Реконструкция облика дна кембрийского моря. Г - голотурии, М - медузы, ГБ - губки, Т - трилобиты. Размер трилобитов - 5-7,5см Некоторое представление о том, как выглядело дно мелкого моря в кембрийское время, дает фото 29. Среди колышущихся морских водорослей находится колония ветвистых губок, напоминающих капусту, голотурии, парашютовидные медузы, а также вымершие ныне организмы, родственные современным ракообразным, - трилобиты. Эта группа живых существ была очень точно воссоздана благодаря тому, что их остатки хорошо сохранились. В одном из районов Британской Колумбии, в самом сердце Скалистых гор Канады, один геолог почти случайно нашел небольшие ископаемые остатки в слое черных сланцев. Осторожно раскопав этот слой, ему удалось найти остатки беспозвоночных среднего кембрийского периода, принадлежавшие к 130 видам; конечности и даже усики животных сохранились до мельчайших деталей (фото 30); они были превращены в уголь, как та рыба, которую мы видели на фото 15. Эта находка представляется исключительной, так как обычно все съедобные части тел мертвых животных на морском дне почти неизбежно уничтожаются трупоядными организмами. В данном случае отложение происходило на дне моря, к которому был прегражден доступ воды, содержащей кислород. В придонных слоях воды, лишенных кислорода, не могли жить какие-либо организмы, кроме некоторых бактерий. Находившиеся здесь организмы погибли, и их тела были погребены на дне, в слое тонкого черного ила. Так как трупоядные организмы тоже не могут существовать без кислорода, то эти тела сохранились до мельчайших деталей, Бескислородные области существуют в морях и океанах и теперь; одна из них находится в Черном море.
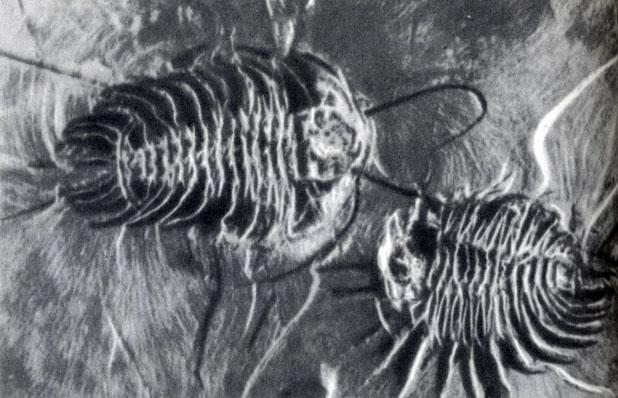
Фото 30. Два ископаемых трилобита (вида Olenoides seratus) в кембрийских черных сланцах Британской Колумбии. Сохранились не только усики, но даже длинные отростки на хвосте Среди 130 видов животных, извлеченных из этого черного сланца, совсем нет плеченогих моллюсков, несмотря на то, что около одной трети кембрийских видов на всей Земле составляли плеченогие. Отсутствие этих животных вряд ли было случайным, Мы вправе предположить, что это связано с местными условиями среды. Плеченогие, которые жили, прикрепляясь к грунту, не обладали подвижностью и не могли плавать. Таким образом, они не могли жить на илистом дне, и, кроме того, они были настолько тяжелы, что не могли быть занесены в эту мертвую зону течениями. То обстоятельство, что на этом небольшом участке морского дна создались условия, благодаря которым сохранились многие ископаемые организмы, необычайно обогатило наши знания о степени развития, достигнутой морскими беспозвоночными в кембрийскую эпоху. Степень этого развития оказалась довольно высокой. Видовой состав кембрийских беспозвоночных резко отличался от современного. Остатки, собранные в кембрийских слоях, содержат в среднем около 60% трилобитов, 30% плеченогих (фото 4) и только 10% других видов. Сейчас трилобиты вымерли, плеченогие обитают лишь на небольших участках океана, а преобладают именно "другие виды". Участок морского дна, изображенный на фото 29, может показаться перенаселенным. Но он не более перенаселен, чем современные коралловые рифы, как может увидеть каждый, кто ныряет с аквалангом. Отбор образцов и подсчет количества ископаемых остатков с каждой единицы объема осадочных пород показывает, что морское дно было повсюду густо населено, возможно, уже задолго до кембрийского времени. Живые существа стараются заполнить все доступное пространство, и если коренные обитатели вытесняются откуда-либо пришлыми организмами, то последние затем также густо заселяют это пространство. Раковины и скелеты Следует отметить одну особенность обстановки, изображенной на фото 29, - присутствие животных, обладающих раковинами и скелетами. Эти приспособления оказали заметное влияние на ход эволюции. Губки имели кремниевые скелеты, а некоторые микроскопические плавающие животные и растения, не показанные на этом рисунке, обладали раковинами из того же материала. В панцирях трилобитов слои фосфата кальция перемежались прослойками хитина - соединения азота, обнаруженного также в ногтях человека; структура панциря напоминала лист фанеры. Большинство многочисленных плеченогих моллюсков и других животных, не изображенных на фото 29, обладали раковинами из карбоната кальция. Раковины - это твердые части тела, которые обычно сохраняются в ископаемом состоянии. Они представляют собой наружный скелет, броню, которая выполняет полезную для животного функцию. Более ранние морские животные имели мягкое тело, у них не было твердых частей, которые могли бы сохраняться. И однако, здесь, в кембрийском море, более полумиллиарда лет назад уже были животные, которые успели обзавестись раковинами трех различных типов.
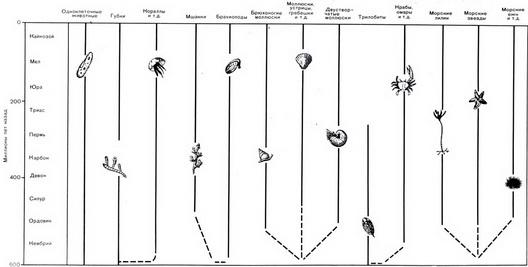
Рис. 40. Основные группы морских беспозвоночных и их положение на геологической шкале. Пунктирными линиями показаны вероятные эволюционные связи между некоторыми группами Палеонтологическая летопись дает нам слишком мало данных, чтобы судить о том, как у морских животных появились твердые части тела, и нам приходится полагаться на теоретические рассуждения. Например, мы можем представить себе, что раковины, состоящие из карбоната кальция, образовались следующим путем. Ионы кальция присутствуют в морской воде и в большей части пищи морских животных. Поэтому кальций мог проникать в тело животного. Большая часть его выводилась из организма, но некоторое количество (зависящее от биохимических особенностей животного) могло оставаться внутри тела или на его поверхности и накапливаться, образуя своего рода затвердение. На морском дне, изобилующем хищниками, вероятно, было безопаснее иметь броню, и эволюция способствовала развитию этих затвердений. В результате постепенно образовались скелеты и раковины. Так это происходило или иначе, но процесс отвердевания частей тела морских животных шел параллельно с процессом развития сосудистой системы у наземных растений. В течение 60 миллионов лет, или около этого, кембрийского периода беспозвоночные с наружным скелетом, имеющим форму раковины, были наиболее сложным видом организмов. Высшие организмы с внутренним скелетом, каким обладают позвоночные животные, в это время еще не появились. Краткий обзор эволюции беспозвоночных Почти все основные группы морских беспозвоночных присутствуют в виде ископаемых в кембрийских слоях. Большая часть из них существует (в измененном виде) и в наши дни. Все данные о развитии этих групп сведены воедино на рисунке 40, В каждой группе происходили определенные эволюционные изменения, но в основном эти изменения были не так резко выражены, как у наземных животных. В большинстве случаев это определяется тем, что в море природные условия менялись в более узких пределах, чем на суше. Тем не менее мы не должны забывать, что корреляция пластов и составление геохронологической шкалы (табл. 1) стали возможны главным образом благодаря наличию различающихся и сходных черт, развивавшихся в процессе эволюции у морских беспозвоночных. Таким образом, чтобы уделить больше внимания историк позвоночных животных, мы ограничимся рассмотрением только двух примеров жизни на мелководном морском дне в различные моменты времени, более поздние, чем изображенный на фото 29. Дно моря в девонское время. Слои песчаников девонского эзраста, выходящие на поверхность в районе Гамильтона в центральной части штата Нью-Йорк, содержат целую коллекцию ископаемых остатков. Образец этих пород показан на фото 31. В центральной части его находится морская звезда Devonaster euchans ("грациозная девонская звезда"), которая очень похожа на современные виды морских звезд. В "северо-восточном" углу образца находится o ее более изящная родственница "хрупкая звезда" - офиура. В "северо-западном" углу, на внешнем образца, находится цилиндрическое углубление окруженное кольцами. Это отпечаток внешней поверхности стебля морской лилии, которая полностью изображена в правой части рисунка 40. Большинство других ископаемых остатков принадлежат трем или четырем видам плеченогих, группе животных, имеющих раковины со створками, как у двустворчатых моллюсков, также изображенных на рисунке 40.
Фото 31. Ископаемые остатки морских беспозвоночных на плите среднедевонского песчанника из окрестностей города Гамильтона, штат Нью-Иорк. Описание см. в тексте. Реконструкция, на фото 32, основана на ископаемых ого возраста, наиденных в данном районе Именно по остаткам фауны этого типа была восстановлена картина, изображенная на фото 32. Ныряя с аквалангом в девонском море, вы могли бы на участке морского дна около 60 сантиметров в поперечнике увидеть нечто подобное изображенному на реконструкции; начальными буквами на рисунке даны названия всех изображенных на нем животных. Особенно следует обратить внимание на двух представителей головоногих одного с длинной прямой раковиной, а другого с волнистой, закрученной. Оба принадлежали к той же самой группе хищников, что и современные кальмары и осьминоги; все они имеют щупальца, снабженные присосками, которые служат для того, чтобы хватать и удерживать добычу. Дно моря в меловое время. На фото 33 представлен другой облик морского дна. Так выглядела в меловом периоде поверхность юго-западной части современного штата Теннесси. Головоногие с прямыми раковинами, которые были широко распространены на более ранних этапах палеозоя, вымерли в ходе эволюции. Вместо них появились моллюски с закрученными раковинами в форме плоских дисков, напоминающих часовую пружину или моток клейкой ленты. В широком устьевом отверстии располагалась голова моллюска, снабженная щупальцами, захватывавшими небольших рыб и другую добычу. Другой особенностью мезозойских морей (в отличие от палеозойских) было присутствие головоногих, подобных кальмарам и носивших название белемнитов. Они имели внутреннюю раковину, напоминавшую спинной хребет и состоявшую из карбоната кальция; большинство этих раковин, сохранившихся в большом количестве в ископаемом состоянии, напоминало толстый карандаш или тонкую сигару. Один из видов белемнитов был гораздо крупнее остальных и имел длину более полутора метров. Белемниты, как и палеозойские головоногие, быстро передвигались, выбрасывая реактивную струю. В основном принцип их движения был тот же, что у современных ракет, если не считать того, что выбрасывалась вода, а не газ. Струя, выбрасываемая головоногими, представляла собой "изобретение" способа движения, совершенно отличного от передвижения с помощью ног или плавников. Кроме того, белемниты, подобно современным кальмарам, имели мешок с чернильной жидкостью, которую они могли выбрасывать через воронку, окрашивая позади себя воду, очевидно, для того, чтобы сбить с толку преследователя. Эти особенности еще раз иллюстрируют своеобразие эволюции в выработке приспособлений к окружающей среде.
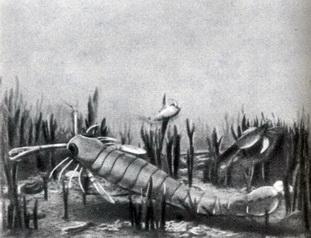
Фото 33. Картина дна мелового моря на месте нынешнего города Мемфис, штат Теннесси. Ширина изображенного участка дна около двух метров. В - водоросли; Б - белемниты; Г - головоногие, с плоскозакрученной раковиной (ГЗ), как у современных наутилусов, с прямой раковиной (ГП) и со спиральной раковиной (ГС); У - улитки "Морские скорпионы". Особо следует упомянуть выделяющуюся своими исключительно крупными размерами группу членистоногих (рис. 40), которые являются дальними родственниками камчатских крабов и, возможно, предками современных скорпионов. Это "морские скорпионы", которые жили в течение большей части палеозоя, но достигли вершины своего развития в морях силурийского времени. Они дышали жабрами, имели сегментную хитиновую оболочку, две пары глаз и шесть пар членистых "ног" (фото 34). Некоторые их виды имели небольшие размеры, но один был более двух метров в длину. Одни виды могли плавать, другие ползали по илистому дну в устьях рек в поисках червей или небольших рыб (фото 35). Превосходя по размерам современных им рыб, "морские скорпионы" были самыми крупными животными, появившимися в биосфере в этот период.
Фото 34. Углефицированный ископаемый 'морской скорпион' (Eusarcus scorpionis) длиной 10 см, найденный в силурийских породах около города Буффало, штат Нью-Йорк. Ясно видны членистое тело, ноги и хвост, кончик которого загнут вбок
Фото 35. Реконструкционный облик самого крупного из известных 'морских скорпионов' длиной более двух метров, а также улиток и трилобитов на дне позднесилурийского эстуария около города Буффало, штат Нью-Йорк. На снимке справа, выше, показан более мелкий скорпион, изображенный в виде ископаемого на фото 34 Насекомые. Как и растения, беспозвоночные вышли из моря на сушу. Наземные моллюски развились из морских моллюсков, насекомые и пауки развились из морских членистоногих, однако об этом мы располагаем до сих пор лишь очень отрывочными сведениями. Из-за малых размеров и отсутствия твердых частей в телах этих животных они могли сохраниться в виде ископаемых лишь в совершенно исключительных условиях. Таким образом, мы мало знаем о борьбе среди растений и животных, переселившихся из моря на сушу. Ископаемые остатки позволяют установить, что скорпионы, возможно, произошли от одного из более мелких видов "морских скорпионов", существовавших еще в силуре, и что немногие насекомые (пока еще бескрылые) и пауки существовали по крайней мере с девона. В то время, как мы знаем, суша уже была покрыта примитивными растениями. До некоторой степени организмы, переселившиеся на сушу, были подготовлены естественным отбором для выработки дальнейших приспособлений к жизни на суше, поскольку морские членистоногие уже имели защитную оболочку и обладали способностью к активному перемещению. Главные изменения, которым они должны были подвергнуться в процессе эволюции для того, чтобы окончательно превратиться в насекомых, - это выработать систему органов дыхания и научиться летать. Первую задачу они решили путем образования многочисленных пор в оболочках; от пор отходили трубки, пронизывающие ткань самого тела. Поэтому у насекомых дыхание и поглощение кислорода. осуществлялось всей поверхностью тела. Вторую задачу насекомые решили к началу позднекаменноугольного времени путем образования выступов оболочки тела, из которых развились крылья. В таком сжатом виде эта история, пожалуй, выглядит повестью об удачливом маленьком членистоногом, которое гналось за "успехами". Но прежде чем нам начнет казаться, что здесь действовали целенаправленные усилия, мы должны вспомнить о факторе времени. Промежуток от среднесилурийского до начала позднекаменноугольного времени составил приблизительно 120 миллионов лет - предположительно в десять раз меньше времени потребовалось для образования Большого каньона реки Колорадо. За это время сменилось по крайней мере 120 миллионов поколений членистоногих; этого времени было вполне достаточно для выработки двух основных приспособлений (для дыхания и полета), которые были "отобраны" из множества попыток и экспериментов. Широко распространенные в позднекаменноугольное время угольные болота, на которых в мелкой воде происходило медленное накопление глинистых отложений, представляли сочетание необычных условий, обеспечивавших сохранность огромного количества насекомых в виде ископаемых. К настоящему времени из них собрано и описано 800 видов только одних тараканов, один из которых достигал 10 сантиметров в длину. Среди других ископаемых насекомых попадаются виды длиной более 30 сантиметров и одна стрекоза с размахом крыльев около 75 сантиметров, почти как у довольно крупного ястреба. Один из этих великанов изображен на фото 26. Очевидно, мягкий климат, благоприятствовавший пышной растительности каменноугольных болот, способствовал также размножению насекомых. Не удивительно, что эти насекомые, только недавно вышедшие из моря и пока еще бескрылые, постепенно приобрели крылья и перешли к воздушному образу жизни. Для мелких живых существ полет в воздухе давал наилучшие возможности спасаться от врагов (от других насекомых, потому что птицы появились только спустя 200 миллионов лет или даже позднее) и преследовать добычу. Необходимо признать, что их образ действия оказался успешным, если под успехом понимать способность заселять территорию и размножаться; доказательством этого служит то обстоятельство, что сейчас число видов насекомых втрое превышает число видов всех остальных групп. Большинство видов насекомых летающие. После того как насекомые поднялись в воздух и приспособились к различным возможным вариантам жизни в воздухе, потребность их в дальнейших изменениях стала ослабевать и ход эволюции замедлился. В начале кайнозоя, после почти 250 миллионов лет развития, многие виды насекомых были уже очень близки к современным. На фото 36 мы видим ископаемое насекомое, которое сохранилось в тонкозернистых отложениях, образовавшихся на дне озера в Центральном Колорадо около 30 миллионов лет назад. Это Glossina, относящаяся к тому же роду, что и современная муха це-це (переносчик сонной болезни), хотя и не к тому же виду. Таким образом, по крайней мере у этого насекомого эволюция происходила довольно медленно.
Фото 36. Ископаемая Glossina (муха це-це) из озерных олигоценовых отложений. Рядом с ней - ископаемый лист дерева, родственного современным березам Происхождение рыб Выше мы описывали особенности формы и строения тела животных, подобных рыбам. И все же остатков организмов, которые могли быть предшественниками рыб, не было обнаружено. Наиболее обоснованное предположение (хотя это всего лишь предположение) состоит в том, что объектом нашего поиска является предок современных морских ежей и звезд, имевших продолговатое мягкое тело, лишенное скелета, не имевший глаз, но обладавший бесчелюстным ртом для всасывания частиц пищи из придонного ила. Одним из оснований для такого предположения послужил вид всех остатков ископаемых рыб, принадлежащих к более ранней эпохе, - силуру (в ордовикских слоях были найдены только костные пластинки). Остатки свидетельствуют о том, что рыбы имели бесчелюстные сосущие рты и не имели плавников. Их тела были одеты довольно гибким покровом из костяных чешуек, а то, что их остатки были обнаружены в отложениях рек и озер, но не морей, доказывает факт переселения этих примитивных рыб вслед за водорослями из морей в реки и озера. На основании указанных фактов можно сделать два заключения. Во-первых, удлиненное тело могло развиться, приспосабливаясь к условиям постоянных речных потоков. Во-вторых, довольно грубые пластинки костяной брони служили не для защиты от хищников (которыми в это время могли быть "морские скорпионы"), а в качестве оболочки, препятствовавшей избыточному проникновению пресной воды через поры внутрь тела. Такая оболочка, вероятно, была необходима для животного, предки которого всегда жили в соленой воде и у которого химизм тела был приспособлен к соленой воде. Таким образом, девонские ископаемые включают морских рыб с челюстями, развившимися из одной из жаберных дуг; они принадлежат к нескольким видам, и самой замечательной из них была хищная рыба из рода Dinictis ("ужасная рыба"), у которой длина тела достигала 9 метров (фото 37). Акулы, изображенные на этом рисунке, обладали хрящевым скелетом, как и современные акулы. Но у Dinictis строение несколько другое, и в ее ископаемом скелете отмечается присутствие кальция. По пути эволюции в этом направлении пошли костистые рыбы, которые также в девонское время развились из более примитивных пресноводных предков. Их скелеты полностью состояли из карбоната кальция и поэтому были более прочными и жесткими. Плавники у них были приспособлены для более быстрого движения в воде, увеличилась и обтекаемость тела. Такие преимущества обеспечили им господствующее положение среди рыб, которое они сохраняют и до сих пор. Мы можем считать их эволюцию успешной, потому что они дали большое количество видов и стали более многочисленными, чем любая другая группа. Но так как почти все эти рыбы продолжали обитать почти исключительно в море, то с точки зрения общего хода эволюции они не получили в дальнейшем стимулов к развитию, как это случилось с вышедшими на сушу животными.
Фото 37. Dinictis, панцирная хищная рыба длиной девять метров, преследующая акул в мелководном девонском море. Во рту у нее вместо зубов выступают заостренные зубчатые края верхней и нижней челюстей Переход к наземной жизни В последней фразе мы сказали "почти все". Исключение составляла одна неприметная группа костистых рыб, которая продолжала оставаться в пресных водах и, таким образом, по существу, находилась на полпути к суше. В результате эти рыбы имели гораздо более интересную историю развцтия, чем их родственники, обитавшие в море. Им пришлось приспосабливаться к изменяющимся условиям жизни на суше, они прошли такой же путь, который до них прошли растения, а затем насекомые, и сумели "порвать" с водной, даже пресноводной, средой и полностью выйти на сушу. Именно они стали предками всех наземных позвоночных животных, в том числе, в конечном счете, и человека. Эти пресноводные представители группы костистых рыб послужили посредниками, через которых человек получил костный скелет, надежно ему служащий. В своих основных чертах скелет, подобный человеческому скелету, развился еще у палеозойских рыб. Рыбы передали нам и основные черты нашего дыхательного аппарата. Произошло это благодаря особенностям наших предков - пресноводных рыб, одной из загадочных групп кистеперых рыб, потомками которых являются современные кистеперые рыбы - целаканты (фото 10). По другой линии развития они явились также предками современных двоякодышащих рыб, которые в наше время обитают в реках сухих областей южного полушария; они имеют вспомогательное легкое, развившееся из плавательного пузыря, с помощью которого могут получать кислород из воздуха. С наступлением сухого сезона двоякодышащие рыбы закапываются в ил на дне речки, образуя подобие кокона, покрытого изнутри слизью. Свернувшись в этом коконе, рыбы впадают в спячку. При этом скорость жизненных процессов составляет лишь Ю % от нормальной во время влажного сезона. При наступлении дождей ил размягчается, рыба высвобождается из кокона и уплывает. Кистеперые рыбы девонского времени имели легкие того же типа. Вначале следует отметить, что в девонских речных отложениях были найдены скелеты Ichthyostega, о чем мы уже упоминали в восьмой главе. Эта рыба принадлежала к амфибиям - группе, название которой означает "ведущие двойную жизнь", то есть частично на суше и частично в воде. К этой группе принадлежат современные саламандры и лягушки. При сравнении амфибии Ichthyostega с девонской кистеперой рыбой видно (рис. 41), что обе эти группы поразительно близки между собой по размерам, форме и строению; особенно велико сходство в строении плавников рыбы и ног амфибии. Еще раз посмотрев на рисунок 31, мы можем провести такое же сравнение между девонской рыбой (кистеперой) и изображенной рядом с ней амфибией. Очевидной, хотя и менее важной, чертой сходства является то обстоятельство, что и у рыбы, и у амфибии голова покрыта броней из костяных пластин. В целом сравнение не оставляет сомнений в том, что девонские амфибии развились из девонских кистеперых рыб.
Рис. 41. Два этапа превращения рыб в наземных позвоночных. А. Девонская кистеперая рыба с примитивным легким. Б. Девонская амфибия Ichthyostega длиной около 90 см. Обратите внимание на лапы с пальцами, все еще соединенными перепонкой, как плавники Безусловно, эти перемены стали возможны благодаря добавочному легкому и особому устройству плавника. Но что явилось толчком к появлению таких перемен? Для объяснения были предложены две правдоподобные гипотезы. Согласно первой, основанной на аналогии с современными двоякодышащими рыбами, причиной явилось изменение климата и появление резко выраженных сухих сезонов, когда реки превращались в разобщенные мелкие водоемы. Оставшись на сухом месте, кистеперая рыба была вынуждена использовать свои необычные плавники для того, чтобы с их помощью перетащить свое тело к ближайшему сохранившемуся водоему, в это время она дышала своим примитивным легким. Экземпляры с менее эффективными плавниками или легкими, или и тем и другим, не могли попасть в соседний водоем и "отбраковывались". Выживали только обладатели наиболее эффективных приспособлений. С течением времени некоторые из выживших рыб стали амфибиями. Другая гипотеза предполагает, что причиной выхода кисте-перых рыб на сушу явилось не высыхание водоемов, а стремление рыб спастись на мелководье от хищников, которых было множество на более глубоких местах. Там они использовали свои плавники сначала для поддержки тела, а затем и для перемещения. Когда в результате естественного отбора плавники достаточно окрепли, рыбы смогли выползать на соседние участки суши, где в девонское время им еще не угрожали хищники и где могло завершиться превращение плавников в примитивные ноги. Конечно, обе эти гипотезы чрезмерно упрощают процесс, но в совокупности, видимо, правильно отражают сущность происшедших перемен. Вспомним, что девонский период длился добрых 50 миллионов лет - время, достаточное для того, чтобы эволюция могла предпринять множество миллионов попыток. Можно было бы с уверенностью предсказать, что за такое большое время какая-то группа озерных или речных животных должна была выйти на прилежащие участки суши. Превращение рыб в амфибий представляет собой заметное достижение на пути эволюции. Так возникла новая династия - четвероногих позвоночных наземных животных. В данной главе мы рассмотрели превращение мягкотелых морских животных в рыб, которым свойственно удлиненное тело, поддерживаемое крепким скелетом, состоящим из множества сочлененных костей, голова с глазами, рот, снабженный челюстями и зубами, сильные плавники и хвост. Обладая всеми этими свойствами, рыба должна была только усовершенствовать легкое, чтобы оно отвечало требованиям среды. После этого процесса она могла полностью выйти на сушу. Литература Мс Аlеster A. L., 1968. The history of life: Prentice-Hall, Inc., Englewood Cliffs, N. J.. p. 39-81. Smith H. W., 1961. From fish to philosopher: The Natural History Library. Anchor Books, Doubleday & Co., Garden City, N. Y., p. 71-101. (Paperback.) Swintоn W. E., 1948. The corridor of life: Jonathan Cape, London, p. 67-122. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||