|
||||
|
|
Глава пятнадцатая. Млекопитающие Как было сказано выше, шкала геологического летосчисления была построена на основе ископаемых остатков. Фанерозой, часть этой шкалы, охарактеризованная многочисленными ископаемыми, которые легко было обнаружить, логически подразделяется на три крупных раздела: палеозой ("эра древней жизни"), мезозой ("эра средней жизни") и кайнозой ("эра новой жизни"). Слово "жизнь" в каждом из этих названий подчеркивает тот факт, что эти подразделения основаны на содержащихся в горных породах ископаемых остатках организмов. В течение кайнозоя господствующими были млекопитающие, мезозоя - рептилии. Основой такого различия является изменение природных условий на континентах и в морях, вызванное главным образом расширением морского дна и движениями континентов. Северная Америка в кайнозойскую эру очень сильно отличалась от того континента, который существовал в мезозое, а изменение условий жизни вызвало эволюцию и возникновение новых форм жизни. Историю млекопитающих лучше всего начать с рассмотрения современных нам млекопитающих, чтобы лучше понять пройденный ими путь эволюции, который привел их к современному состоянию. Мы уже говорили, что характерными особенностями млекопитающих являются молочные железы для вскармливания детенышей, теплая кровь (что фактически означает постоянную температуру тела днем и ночью), волосяной покров, служащий для термоизоляции, четырехкамерное сердце, дифференцированные зубы, служащие для откусывания и жевания, и хорошо развитый мозг. Эти черты общие для всех млекопитающих. По особенностям же процесса размножения млекопитающие подразделяются на три группы. 1. Наиболее примитивные - однопроходные, потому что они откладывают яйца (хотя и вскармливают детенышей молоком); они отличаются также некоторыми особенностями анатомии, которые в ископаемых остатках не устанавливаются. Сейчас их существует только два рода - утконос с клювом, как у утки, и австралийская ехидна (Отряд однопроходные объединяет два семейства: Tachyglossidae (ехидны) и Ornithorhynchidae (утконосы). В составе семейства ехидны - два рода, утконосов - один род. - Прим. ред). Оба эти рода очень хорошо приспособлены к определенному образу жизни и ограничены в распространении Австралией, где, по-видимому, конкуренция слабее, чем в других частях света, что и создает предпосылки для выживания самых примитивных животных. Поскольку ископаемые однопроходные встречаются только в Австралии, возможно, что в других местах они никогда не существовали. 2. Промежуточную группу образуют сумчатые, из которых, пожалуй, наиболее общеизвестны кенгуру. Детеныши у кенгуру рождаются живыми, они очень слабо развиты; новорожденный кенгуру гложет иметь длину до трех сантиметров. Детеныши заползают в сумку матери и в течение долгого времени питаются там молоком, пока не вырастут в несколько раз больше (по сравнению с размером, который они имели при рождении) и смогут выйти из сумки и кормиться самостоятельно. Сейчас сумчатые живут главным образом в Австралии; к ним относятся кенгуру, "медведи" коала и вомбаты. Единственное сумчатое, сохранившееся в Северной Америке, - опоссум, - по-видимому, приспособилось к конкуренции, живя на деревьях. Размножение у сумчатых более безопасно, чем откладывание яиц, потому что развивающиеся детеныш постоянно находится под защитой матери. Но еще более усовершенствовала этот процесс третья группа млекопитающих. 3. Наиболее развиты плацентарные млекопитающие, названные так потому, что у них есть плацента - орган, находящийся внутри матки и напоминающий мембрану, пронизанную многочисленными кровеносными сосудами. Через плаценту эмбрион получает питательные вещества и кислород, и через нее же происходит удаление углекислого газа и отходов. Все это устройство (рис. 53) напоминает амниотическое яйцо (рис. 43), от которого оно, очевидно, и произошло. Но плацента дает млекопитающим преимущества не только по сравнению с яйцекладущими животными, но и по сравнению с сумчатыми. Эмбрион, развитие которого регулируется плацентой, может находиться в матке до своего полного и окончательного развития. Детеныш плацентарных млекопитающих, по крайней мере у большинства родов, после того, как он родится, довольно скоро может заботиться сам о себе. Плацента внесла существенное усовершенствование в процесс размножения.
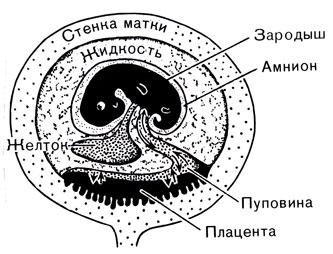
Рис. 53. Основные черты зародыша и матки у плацентарных млекопитающих (сравните с рис. 43). На ранних стадиях зародыш питается желтком, который затем исчезает. Зародыш получает питательные вещества и кислород и выделяет углекислый газ и отходы через плаценту. Направление этого обмена показано стрелками Мезозойские млекопитающие С нашей "позднекайнозойской" точки зрения, современные млекопитающие находятся почти в конце эволюционной линии. Рептилии пермского и триасового периодов, которые походили на млекопитающих, находились на другом конце эволюционной линии. Необходимо помнить, что путь развития организмов от наиболее примитивных млекопитающих до человека не был прямым и это развитие происходило не всегда с одинаковой скоростью. В течение почти 250 миллионов лет мезозоя эволюция млекопитающих не представляла ничего интересного. Можно сказать, что они выжидали своего часа. В то же самое время настоящие рептилии распространились очень широко и приспособились к различным условиям среды в мягком климате. Низменный рельеф и мягкий климат мезозоя способствовали развитию рептилий, но не представляли исключительно благоприятных условий для млекопитающих. Вновь появившиеся млекопитающие должны были выжидать, пока не уменьшились численно и не вымерли динозавры и пока в кайнозое не возникли новые типы природной среды, лишь после чего они в свою очередь смогли сильно распространиться и занять господствующее положение. Все это ясно говорит нам, что эволюционное развитие меняет свое направление в соответствии с меняющимися возможностями, которые предоставляет окружающая среда. Пока происходит движение плит коры и перемещение континентов в более высокие или более низкие широты, а иногда и соединение континентов, будет происходить и изменение условий жизни, создающее новые возможности для эволюции живых организмов. Вполне вероятно, что предком настоящих млекопитающих мог быть Cynognathus (фото 42) или один из родственных ему видов. А может быть, эта честь принадлежала другой рептилии, еще не обнаруженной в ископаемом состоянии. Это не имеет большого значения, потому что млекопитающие и рептилии, подобные млекопитающим, очень тесно связаны между собой. Какой бы ни была рептилия - родоначальник млекопитающих, - произошедшие от нее млекопитающие имели очень небольшие размеры. Большинство ископаемых млекопитающих, обнаруженных в триасовых, юрских и меловых слоях, были размером с крысу или мышь, а самые крупные не больше кошки. До сих пор не найдено ни одного целого черепа, не говоря уже о скелете, но обнаружено множество мелких челюстей с дифференцированными, как у млекопитающих, зубами и разрозненные кости. Эти кости больше напоминают кости сумчатых или однопроходных, чем плацентарных млекопитающих. В меловых слоях в западной Канаде найдены остатки, принадлежащие, без сомнения, опоссуму. Небольшие размеры мезозойских млекопитающих указывают на то, что они не могли конкурировать с динозаврами и другими рептилиями, несмотря на имеющийся у них сравнительно большой мозг. Можно предполагать, что млекопитающие занимали подчиненное положение, старались держаться в стороне и, вероятно, питались насекомыми и семенами растений. Так как мы не располагаем целым скелетом, мы не можем представить себе, как они выглядели, но ископаемые черепа позволяют воссоздать внешний вид головы этих млекопитающих (рис. 54).
Рис. 54. Голова позднемезозойского млекопитающего. Остальные части тела реконструировать пока не удается, так как найден только череп Приспособления к кайнозойским природным условиям Общий обзор. Таково было положение вещей в конце мезозоя. На сцене начали появляться млекопитающие. С изменением ландшафтов и похолоданием климата динозавры постепенно исчезали, одна группа за другой, оставляя жизненное пространство свободным. Млекопитающие, которым больше не угрожали рептилии, осмелели. Волосяной покров и теплая кровь, а также больший объем мозга давали им возможность выжить в условиях похолодания и приспособиться к новым, более разнообразным природным условиям, на что динозавры оказались неспособны. По мере того как рептилии сдавали свои позиции, млекопитающие занимали освободившиеся территории, где им не угрожала существенная конкуренция. Наконец рептилии исчезли. Единственной опасностью для этих первых млекопитающих были другие млекопитающие. Конечно, среди "других млекопитающих" вскоре появились плотоядные хищники, которые завоевали свое место в пищевой цепи. В этой истории можно найти сходство с мезозойской историей рептилий. Новая форма распространяется на новые территории и там, встречаясь с различными условиями среды, которую можно заселить, разделяется на несколько разновидностей. Животные одного вида охотятся на других. В результате конкуренции устанавливается своего рода динамическое равновесие видов. Оно "динамично" потому, что при этом одни виды вымирают, другие развиваются. Это равновесие представляет собой бесконечный ряд "битв", в которых перевес получает то одна, то другая сторона. Если кайнозойскую историю млекопитающих рассматривать в перспективе, она состоит как бы из двух фаз: ранняя фаза экспериментирования природы на млекопитающих и поздняя фаза отработки их деталей, во время которой направления эволюции становятся более отчетливо выраженными. Ранняя фаза продолжалась включительно по олигоцен, поздняя - остальную часть кайнозоя. Конечно, она продолжается и сейчас, но с тех пор, как в конце плиоцена господствующее положение занял человек, путь развития млекопитающих резко изменил свое направление. Палеонтологическая летопись ранней фазы создает представление, что эволюция довольно часто экспериментировала, временами неудачно, разрабатывала и опробовала различные модели, выбирая те, которые могли бы выдержать конкуренцию и преуспеть в той или иной природной среде. Во многих ранних моделях были обнаружены недостатки, и они были одна за другой отвергнуты. Уцелевшие виды просуществовали всю раннюю фазу и в ходе поздней фазы оформились и развивались по вполне определенным направлениям. Многие из них существуют до сих пор, занимая господствующее положение среди млекопитающих новейшего времени.
Фото 48. Uintatherium, травоядное млекопитающее массивного сложения, высотой около 1,5 м на фоне реконструированного эоценового ландшафта. Его причудливо украшенный череп едва ли был полезен при обороне или при нападении. Возможно, он привлекал особей противоположного пола, а если так, то естественный отбор мог способствовать развитию этих выростов Ранние неспециализированные группы. Примитивный характер раннекайнозойских млекопитающих (фото 48, рис. 55) объясняется двумя главными причинами. Во-первых, эти млекопитающие происходили непосредственно от позднемеловых видов, которые сами по себе были примитивными. Во-вторых, широко распространенные в это время лесные ландшафты привели к сохранению архаичных форм животных. Вероятно, большая часть раннекайнозойских млекопитающих принадлежала к группе плацентарных. Как правило, они были небольших или средних размеров (хотя один из них достигал величины современного носорога). Ступня этих млекопитающих была плоской или почти плоской (рис. 55, А), а у некоторых массивной, как у слона, и почти все они имели пять пальцев. Среди них были как травоядные (большей частью питающиеся побегами деревьев), так и хищники. Последние выделялись главным образом формой зубов и примитивными когтями, не имея резких отличительных особенностей общего облика. Некоторые из раннекайнозойских млекопитающих, как, например, Uintatherium, показанный на фото 48, обладали странными костными выростами на черепе, а самцы этого вида имели также длинные верхние клыки.
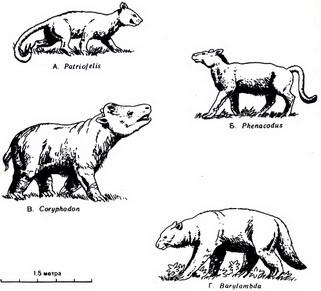
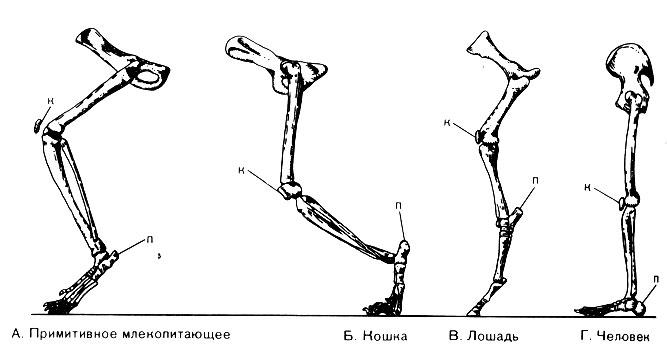
Рис. 55. Четыре представителя ранних и примитивных кайнозойских млекопитающих. A. Patriofelis, хищник. Б. Phenacodus, небольшое травоядное. Один из его родственников, возможно, был родоначальником линии лошадей. В. Coryphodon, травоядное массивного сложения, возможно, обитавшее в болотах. Г. Barylambda, травоядное длиной около 2,5 м Среда обитания и пища. Один за другим примитивные виды млекопитающих вымирали или в ходе эволюции становились более развитыми формами при определяющем влиянии двух главных факторов - окружающей среды и пищи. В раннем кайнозое были широко распространены леса, редколесья и саванны. В таких условиях листья деревьев и кустарников составляли основную пищу растительноядных животных, медленно передвигающихся от дерева к дереву. От врагов эти животные спасались главным образом за счет "маскировки", поскольку большинство из них не могло бегать на большие расстояния. Здесь встречались и хищники, и они также прибегали к маскировке, выслеживая добычу. Длительного преследования травоядных животных хищниками обычно не происходило. Напротив, в степях, которые начали появляться в эоценовом периоде и все шире распространялись в остальную часть кайнозоя, травоядные могли питаться только травой и злаками. Такие млекопитающие должны были наклоняться за пищей к самой поверхности земли. Им приходилось постоянно двигаться в поисках свежей травы и воды и, таким образом, перемещаться на далекие расстояния. Им также приходилось спасаться бегством от неизбежных хищников, так как спрятаться здесь было негде. Их шансы на выживание могли увеличиться лишь за счет усиленного размножения или объединения в большие группы или стада. Перед хищниками вставала необходимость "развития" большой скорости, чтобы преследовать добычу, так как они также не могли маскироваться, выслеживая добычу. Мы знаем, что некоторые млекопитающие всеядны. Например, современные медведи едят плоды, ягоды, муравьев и рыбу. Они могут существовать в различных условиях, обычно там, где есть реки и по крайней мере какие-нибудь деревья, поскольку у них мало врагов и скорость передвижения не имеет для них большого значения. Мы вправе ожидать, что такие различия в условиях жизни млекопитающих и пищевом рационе существенно повлияли на их форму тела и строение. Это становится ясно, когда мы изучаем ряд ископаемых скелетов, полученных из достаточно полного геологического разреза. Эволюция конечностей и зубов. Ископаемые скелеты, собранные в различных отложениях мезозоя, обнаруживают прогрессирующие изменения в строении, очевидно связанные со средой обитания и с тем способом питания, который был обусловлен этой средой. Млекопитающие, которые паслись в лесах и редколесьях, сохраняли примитивную, довольно плоскую стопу, которая, как мы предполагаем, была типична для мезозойских млекопитающих. У них сохранялись также неспециализированное коренные зубы, служившие для пережевывания лиственной пищи. Постепенно ноги их удлинялись, что отчасти позволяло им лучше спасаться от хищников, которые постепенно становились более опасными. У плотоядных животных, которые охотились и выслеживали травоядных, также сначала сохранялась плоская стопа. Но понемногу их конечности приспособились к быстрому бегу на короткие расстояния. Пятка (п) у них отделилась от земли (рис. 56, Б), конечности стали длиннее и легче, и мускулы бедра образовали пучок, необходимый, чтобы обеспечить быстрый рывок в начале преследования. Хорошо развитые когти позволяли схватывать и валить добычу. Заостренные передние зубы давали возможность кусать, а сильные мышцы челюстей грызть добычу.
Рис. 56. Кости конечностей четырех различных видов млекопитающих Конечности и зубы травоядных млекопитающих также претерпевали изменения, хотя совсем иначе. Так как пасущиеся животные должны были бегать, у них значительно увеличилась длина и бедра и стопы; мускулы стали более сильными. Пятка приподнялась так высоко, что стопа стала почти вертикально (рис. 56, В), а когти на пальцах стали длиннее и толще. Параллельно с этим произошло удлинение отдельных пальцев, так что другие перестали прикасаться к земле, постепенно уменьшились и в конце концов совсем исчезли (рис. 57). Такое изменение уменьшило вес стопы и способствовало увеличению скорости бега у травоядных млекопитающих и дальности их перемещения. Эта эволюция, происходящая более или менее одновременно у различных видов млекопитающих, может служить свидетельством определяющего влияния среды. Однако вследствие дифференциации среды изменение стопы зашло у отдельных видов млекопитающих дальше, чем у других. У свиней "действуют" четыре пальца, у носорогов - три, у верблюдов, оленей и крупного рогатого скота - два, у лошадей - один.
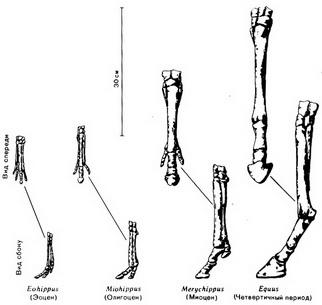
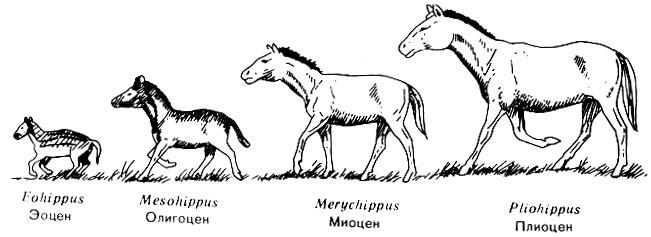
Рис. 57. Кости передних конечностей четырех родов ископаемых лошадей. Через 50 миллионов лет эволюционного развития число пальцев уменьшилось от четырех до одного, а общая длина конечности увеличилась почти в 12 раз Этот процесс занял долгое время. Например, лошади, впервые появившиеся в эоцене, дошли до своего современного состояния (то есть стали иметь один палец) к раннему плиоцену, а это значит в течение почти 60 миллионов лет. Если поколения лошадей сменяются в среднем через четыре года, то этот процесс охватил 15 миллионов поколений - 15 миллионов возможностей для изменений, причем каждое поколение подвергалось воздействию среды, имевшему то же самое направление развития. Если бы мы взяли 15 миллионов лошадей, по одной от каждого поколения, и выстроили их друг за другом, начиная с небольшой эоценовой лошади и кончая крупной современной, и они проходили бы мимо нас со скоростью около 6,5 километра в час, то этот парад продолжался бы более четырех месяцев. Наблюдая эту процессию в течение часа (в любой момент из этих четырех месяцев), мы не увидели бы заметных изменений в облике животных. Но к концу процессии вместо небольших, питающихся ветками животных высотой около 30-40 сантиметров, передвигающихся, опираясь на четыре пальца, мы увидели бы лошадей, опирающихся на один палец и имеющих высоту в холке до полутора метров (рис. 58).
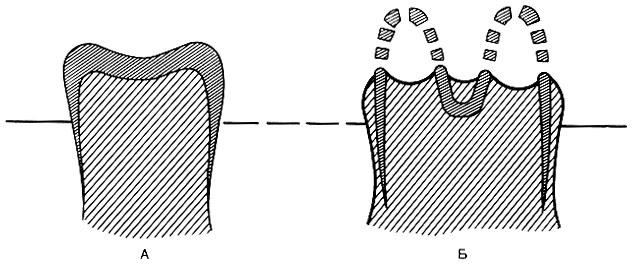
Рис. 58. Четыре вида лошадей, живших в различные эпохи. Eohippus и Mesohippus в действительности были еще меньше по отношению к более поздним лошадям, чем это показано на рисунке В ископаемых скелетах степных травоядных заметны и другие эволюционные изменения. У них удлиняются челюсти, причем происходит разделение зубов на резцы, расположенные спереди, и зубы, расположенные сзади и предназначенные для жевания. Но так как степные травы содержат большое количество кремния, а кроме того растительность степи часто покрыта пылью, серьезную проблему представляло истирание коренных зубов мелкими твердыми частицами кремнезема. Ход эволюции привел к видоизменению коренных зубов. У лошадей эти зубы приобрели высокую коронку и складчатые образования эмали (рис. 59), напоминающие модель Аппалачских гор в миниатюре. Эти складки включают твердый слой эмали и прилегающий к ней более мягкий материал. При жевании пыльной травы зубы снашиваются, но эмаль стирается в последнюю очередь и образует выступы, придавая поверхности зуба шероховатость и облегчая перетирание зеленой пищи.
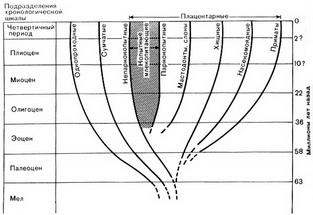
Рис. 59. Коренные зубы млекопитающих. А. Низкий коренной зуб нетравоядного млекопитающего. Б. Коренной зуб травоядного млекопитающего, более высокий и имеющий складчатую структуру; более мягкий материал быстрее истирается, а твердая эмаль (на рисунке показана частой штриховкой) образует небольшие выступы В отличие от описанных случаев далеко зашедшей специализации, некоторые млекопитающие были очень слабо специализированы. Они не были ни травоядными, ни плотоядными, могли есть и действительно ели все, что попадалось. Они не оказывали четкого предпочтения ни лесу, ни степи, но жили в любом районе, где была хорошая пища в достаточном количестве, будь то насекомые, ягоды или плоды, рыба или мелкие млекопитающие. Они были искателями и скитальцами. Типичными современными примерами таких животных являются медведи, еноты, некоторые виды обезьян. Млекопитающим, ведущим такой образ жизни, не требовалось вследствие условий жизни или пищевого рациона изменять форму своих зубов и конечностей. Они навсегда сохранили примитивную плоскую стопу (рис. 56, Г) и низкие коренные зубы (рис. 59 А). Эволюция млекопитающих на суше, в море и в воздухе Если проследить эволюционные линии всех ископаемых млекопитающих, как мы это уже сделали с лошадьми, эти линии, разветвляясь, образуют дерево, подобное схеме развития динозавров. Схема млекопитающих значительно сложнее, но на рисунке 60 дан ее упрощенный вариант, включающий только основные ветви. Однако даже на этой упрощенной схеме видно, что большинство ветвей отделилось от главного ствола в эоцене. История эволюционных изменений основных линий слишком сложна, чтобы излагать ее полностью. Интересно, однако, рассмотреть хотя бы немногие из исчезнувших ныне видов, принадлежавших к заканчивающимся тупиком эволюционным линиям, не показанным на нашей схеме. Некоторые из них изображены на рисунке 61. С нашей точки зрения, они выглядят странно. Но было бы, однако, несправедливо считать эти виды неудачными только на том основании, что они вымерли и, поэтому их облик непривычен для нас.
Рис. 60. Эволюционные линии млекопитающих В отличие от странных животных, показанных на этом рисунке, крупные хищники сохраняли относительную стабильность, не подвергаясь таким сильным эволюционным изменениям, как лошади. В начале кайнозоя, как было установлено, кошки подразделились на две группы - кошки, к которым относятся львы, тигры и современные домашние кошки, и саблезубые кошки более массивного сложения. У последних (фото 49) верхние клыки были длинными и уплощенными, а челюсти видоизменены таким образом, что могли широко раскрываться. Прыгая на добычу, такая кошка глубоко вонзала свои саблеобразные клыки, а затем вытаскивала их, оставляя глубокие кровоточащие раны.
Рис. 61. Некоторые млекопитающие, обладавшие приспособлениями, оказавшимися неудачными. A. Platybelodon, млекопитающее, похожее на слона, у которого рот напоминал огромный совок; им животное сгребало болотные растения. Миоцен. Б. Dinotherium размером со слона, имел загнутую книзу нижнюю челюсть с клыками. Миоцен.
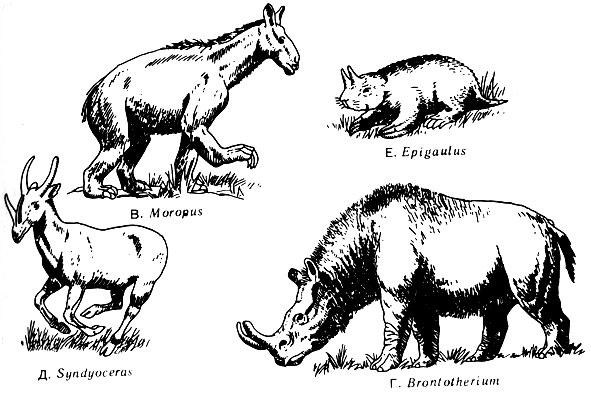
Рис. 61. Некоторые млекопитающие, обладавшие приспособлениями, оказавшимися неудачными. В. Moropus, похожий на лошадь, но более крупный, - травоядное животное с короткими задними ногами. Его трехпалая стопа была снабжена когтями, возможно, служившими для выкапывания растений. Миоцен. Г. Brontotherium, травоядное длиной более 3,5 м. Олигоцен. Д. Syndyoceras, родственный оленю, но с дополнительным рогом. Миоцен. Е. Epigaulus, рогатый грызун с необычайно длинными когтями. Плиоцен К олигоцену обе группы кошек так хорошо приспособились к своему хищному образу жизни, что с тех пор они мало менялись. Саблезубый тигр вымер немногим более 10 000 лет назад, но мы не можем считать его неудачной моделью только на том основании, что он вымер. Два или более видов его существовали на протяжении более 30 миллионов лет. И мы не можем считать, что здесь эволюция зашла в тупик. Действительно, вполне может быть, что саблезубые исчезли только потому, что человек каменного века со своими усовершенствованными орудиями охоты успешно боролся с ними за пищу. Как мы уже видели, кайнозойские млекопитающие повторили ход развития рептилий - подобно им, они увеличивали разнообразие видов и численность и достигли господствующего положения. Как и мезозойские рептилии, они успешно проникли в водную и воздушную среду. У мелких млекопитающих, живущих на деревьях, развились крылья, состоящие из кожистой перепонки, и тем самым было положено начало эволюционной линии летучих мышей. Крыло летучей мыши сконструировано лучше, чем крыло летающей рептилии (рис. 52), но оно уступает крылу птицы по структуре, а также потому, что на нем нет перьев. Может быть, это и есть главная причина того, почему летучие мыши никогда не достигли такого успеха, как птицы.
Фото 49. Сцена времен олигоцена на территории западной части современных Соединенных Штатов; саблезубая кошка (Hoplophoneus) убивает примитивную лошадь (Mesohippus). Реконструкция В водной среде млекопитающие также повторили путь развивающейся группы мезозойских рептилий. Не успела начаться эоценовая эпоха, как с суши в море переселились наземные хищники, превратившиеся в примитивных китов с острыми зубами. Возможно, они прошли промежуточную стадию, заходя на мелководье и охотясь на рыбу. Один из этих видов развился позднее в настоящих китов, которые разделились на несколько известных сейчас видов. Самое крупное из ныне живущих морских млекопитающих, синий кит, по длине равен самому большому из когда-либо существовавших динозавров, а по весу превосходит его более чем вдвое. В точности так же, как это произошло у их предшественников - рептилий за 200 миллионов лет до них, у всех морских млекопитающих тело удлинилось и конечности превратились в плавники или ласты. Снова мы с поразительной ясностью видим способность среды изменять форму и строение организмов. Притягательная сила кайнозойских морей действовала не только на китов. В эоцене одно из растительноядных млекопитающих, возможно состоявшее в отдаленном родстве с предками слонов, также погрузилось в воду и присоединилось к обитателям моря, основав линию, которая привела к современным морским коровам и ламантинам. И в довершение к этому в миоцене хищник, предположительно родственный кошачьим, присоединился к прежним обитателям моря и дал линию современных тюленей. Дальние миграции Палеонтологическая летопись ясно свидетельствует о том, что млекопитающие неоднократно мигрировали с одного континента на другой. Мы находим один и тот же род или близкие роды на двух разобщенных массивах суши. Как могло произойти такое расселение? Пыльца растений переносится по воздуху на тысячи километров, но пылинки имеют микроскопические размеры. Насекомые заносятся ветром за сотни миль водных пространств. Семена растений могут переноситься течениями через Атлантический океан. Стволы деревьев также проплывают большие расстояния. Известны случаи, когда змеи и крокодилы (и те и другие привычны к водной среде) проплывали сотни километров между отдельными островами. Полярные медведи проходят сотни километров по поверхности морских льдов. Но все это частные случаи, которые не могут объяснить данные палеонтологической летописи, свидетельствующие о том, что десятки видов млекопитающих размерами от мыши до слона переходили с одного континента на другой. Такие миграции нельзя объяснить с помощью гипотезы движения плит коры, при котором происходит разъединение континентов и отторжение их частей далеко друг от друга, потому что Атлантический океан существовал задолго до начала кайнозоя. Что же касается возможности переплыть морское пространство, то очень мелкие млекопитающие, возможно, и могли переплывать его на стволах деревьев, но для большинства остальных животных единственный путь с одного континента на другой (если не пользоваться целой флотилией ковчегов гораздо большего размера, чем знаменитый ковчег Ноя) - сухопутные мосты. Обратимся к карте континентов (рис. 16), на которой показаны также шельфы. На этой карте, во-первых, видны узкие перемычки суши, существующие сейчас между двумя Америками и между Европой и Африкой, и, во-вторых, мелководные участки, где при довольно незначительных изменениях уровня океана относительно суши два или более массивов суши могут оказаться соединенными (или почти соединенными цепочкой островов между ними). Примером может служить связь Евразии с Северной Америкой через Сибирь и Аляску и Евразии с Австралией через острова Малайского архипелага. Таким образом, при небольших подвижках суши или изменениях уровня моря могут возникнуть или мосты суши, или мелководья, соединяющие между собой все континенты, кроме Антарктиды (отметим, что речь идет лишь о последних 63 миллионах лет). Мост между Аляской и Сибирью. Перемычка суши между Аляской и Сибирью может быть восстановлена, если суша поднимется или уровень моря снизится всего лишь на 45 метров, Есть основания полагать, что здесь в течение большей части кайнозоя существовал широкий мост суши. По нему происходило перемещение различных млекопитающих в обоих направлениях. Вот почему среди ископаемых млекопитающих Америки и Евразии так много близких форм и почему некоторые из них почти совершенно идентичны. Мы, конечно, не можем считать, что эти млекопитающие обладали таким "интеллектом", что могли "запланировать" и осуществить это длинное путешествие в одну сторону. Они просто перемещались в направлении, где была хорошая, удовлетворявшая их пища и где было не слишком много конкурентов, претендующих на нее. Паразитирующие насекомые переселялись вместе с травоядными животными, а крупные кошки и другие плотоядные играли роль убийц и грабителей на большой дороге. Две Америки. Иной была история Панамского перешейка, соединяющего сейчас две Америки; она также оказала сильное влияние на развитие млекопитающих в кайнозое. В палеоцене и раннем эоцене перешеек был сушей. Это позволило некоторому количеству очень примитивных, неспециализированных видов просочиться на южный континент. Затем позднее, в эоцене, связь между континентами была прервана в результате опускания перешейка под воду; он оставался погруженным в течение почти 50 миллионов лет, до конца плиоцена. В течение этого времени примитивные млекопитающие (фото 50), в том числе сумчатые, перекочевавшие в Южную Америку до того, как прервалось сообщение между континентами, медленно развивались, защищенные от вторжения более развитых видов из других мест. Но когда мост был восстановлен, огромное количество развитых, современных млекопитающих (включая крупную кошку, называемую ягуаром) нахлынуло сюда с севера и стало конкурировать с более примитивными коренными обитателями. Один за другим местные виды исчезали, и у нас есть основания полагать, что своей печальной участью они были в значительной степени обязаны также первобытному человеку.
Фото 50. Необычайные южноамериканские млекопитающие, обитавшие на территории Аргентины в плейстоцене. Слева - наземные ленивцы, справа - глиптодонты (гигантские броненосцы) с панцирем, как у черепах. Реконструкция Как и на мосту, соединявшем Аляску и Сибирь, движение с одного американского континента на другой происходило по двум направлениям, но поток, направленный на север, был не столь уж велик. В Северную Америку проникли броненосцы, наземные ленивцы и глиптодонты (гигантские броненосцы), причем первые из них до сих пор обитают в Техасе. Сумчатые Австралии. Каждый знает кенгуру - этот национальный символ Австралии. Фактически, как и все другие млекопитающие Австралии, кенгуру относятся к сумчатым млекопитающим. Благодаря своему примитивному строению они не могут конкурировать с плацентарными млекопитающими, и, если бы последние смогли появиться в Австралии в большом количестве, они, вероятно, давным-давно вытеснили бы сумчатых. В Северной Америке, где сумчатые и плацентарные одно время конкурировали между собой, все процветавшие ранее сумчатые исчезли, за исключением представителей одной ветви, которая привела к опоссуму, единственному сохранившемуся здесь до сих пор сумчатому. В Австралии сумчатые жили гораздо лучше, и причиной этого была слабая конкуренция со стороны других наземных животных. Среди современных животных Австралии, помимо сумчатых, встречается только два вида однопроходных, несколько плацентарных грызунов, похожих на крыс, летучие мыши, мелкие рептилии и амфибии. Почти полное отсутствие плацентарных млекопитающих среди местной фауны лучше всего объясняется изложенной ниже гипотезой, которая хотя и не доказана, но очень правдоподобна. На карте (рис. 16) можно видеть, что между Австралией и соседней с ней Азией существует обширное мелководье, на котором так много островов, что проливы между ними нешироки. Предполагается, что в самом конце мелового периода или в начале палеоцена примитивные сумчатые попали в Австралию, "прыгая" с острова на остров, то есть отдельные особи так или иначе случайно преодолевали узкие проливы между островами. Они заселяли один остров, затем происходило новое случайное переселение, и так продолжалось до тех пор, пока сумчатые не достигли Австралии. В течение 20 или 30 миллионов лет ряд таких происшествий должен был произойти неизбежно. Если мы примем предположение, что австралийские сумчатые произошли от этих путешественников, перебиравшихся с острова на остров, мы должны принять также и гипотезу, что плацентарные млекопитающие могли осуществить это так же, как и сумчатые, если не лучше. Возможно, что, когда Австралия в какой-то момент до начала мелового периода отделилась от предположительно существовавшего тогда древнего большого континента, показанного на рисунке 27, и стала перемещаться к северу вместе с плитой земной коры, на ней уже жили ранние формы сумчатых. Плацентарные млекопитающие в это время еще не появились. Но какое бы объяснение мы ни приняли, результат остается одним: сумчатые в Австралии были изолированы. Поэтому их развитие могло идти различными путями, избегая губительной конкуренции со стороны более удачливых плацентарных млекопитающих. Развитие сумчатых в основном заключалось в приспособлении к различным типам природной среды, встречающимся в пределах обширного австралийского континента. В пределах каждого типа среды сумчатые приобретали такие формы, которые были лучше приспособлены к данному образу жизни, - формы, напоминающие плацентарных млекопитающих, живущих в подобных условиях. Так, например, в число современных сумчатых входят вомбат, бандикут, сумчатая летяга, сумчатый (тасманийский) волк и сумчатая мышь. Эти животные могут быть сопоставлены соответственно с плацентарными североамериканским сурком, кроликом, белкой-летягой, волком и мышью. Эти сопоставления еще раз показывают сильное влияние условий окружающей среды на ход эволюции; форма тела животного и его способности вырабатываются в соответствии с образом жизни, обусловленным внешними условиями, вне зависимости от того, каким был исходный материал эволюции. Позднее, уже в кайнозое, появились плацентарные животные, напоминающие крыс, а за ними летучие мыши и змеи. Летучие мыши легко могли быть занесены с ветром во время штормов; остальные могли приплыть на стволах деревьев. Они не представляли сколько-нибудь серьезной угрозы с точки зрения борьбы за существование и, как кажется, не повлияли сколько-нибудь существенно на развитие сумчатых. Жизни сумчатых ничто не угрожало до тех пор, пока сравнительно недавно не появился человек - охотник, и с ним другой охотник - собака. Прибытие европейцев в Австралию имело катастрофические последствия. Сейчас сумчатые один за другим исчезают. На примере одного из животных, которых завез человек, - кролика (плацентарного) - можно видеть, как происходит расселение новых видов животных на свободной территории. В 1859 г. из Англии в Австралию были завезены две дюжины кроликов. В течение следующих 69 лет потомство этих кроликов распространилось почти по всей Австралии. На площади, составляющей приблизительно 1/3 всей страны, в 1928 г. был сделан подсчет, показавший, что кроликов здесь приблизительно полмиллиарда. Эта цифра помогает понять, как шло расселение рептилий в мезозое. Заключение. Таким образом, расселение наземных животных во всем мире очень близко к тому, какое должно было бы быть, если бы существовали перемычки суши или цепочки островов между северными континентами и не существовало никаких мостов, по крайней мере в кайнозое, к югу от Австралии или Южной Америки. Эти два континента представляли собой тупики, потому что на них обитают в основном животные, принадлежащие к тупиковым линиям эволюции. Временами, на протяжении 50 миллионов лет или более, на них попадали млекопитающие, но дальше они двигаться не могли. Мостов суши, соединявших их с Антарктидой, не существовало. Благодаря отдаленности и низким температурам антарктического континента животные попадали сюда только изредка и случайно. Даже сейчас высшие животные представлены там только тюленями и пингвинами; и те и другие могли попасть сюда и без всяких мостов. Значение истории млекопитающих Кайнозойская история млекопитающих далеко не такая длинная, как история мезозойских рептилий, но для нас она гораздо важнее. Она прослежена на основе гораздо большего количества ископаемых и представляет более разнообразные виды животных. География и природные условия кайнозоя гораздо ближе к современным, чем мезозойские. Таким образом, нам более знакома обстановка, в которой развивались события. Наконец, млекопитающие принадлежат к той же группе, что и мы сами, и потому представляют для нас большой интерес. В развитии млекопитающих мы более ясно видим возникновение особенностей строения и функций, которые существуют и в нашем теле. В этой главе мы не останавливались на структурных и функциональных особенностях организмов; мы даже не упомянули приматов, особую группу, к которой принадлежим и мы сами, ограничившись указанием ее места в общей системе на рисунке 60. Приматы для нас настолько важны, что они заслуживают особого рассмотрения. После того как мы рассмотрим природные условия позднего кайнозоя и климатическую обстат новку, создавшую эти условия, мы обратимся в главе восемнадцатой к приматам и посмотрим, к чему они пришли в результате своего развития. Литература Fentоn С. L., Fеntоn M. A., The fossil book: Doubleday and Co.. Inc. Garden City. N. J.. p. 375-435. Rоmer A. S., 1959, The vertebrate story: University ot Chicago Press, p. 219-308. Scott W. В., 1937. History of the land mammals in the western hemisphere: The Macmilan Co., New York. |
|
||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Другие сайты | Наверх |
||||
|
|
||||